



2. 三沙美济渔业开发有限公司 三亚 572000;
3. 上海海洋大学水产与生命学院 上海 201306
2. Sansha Meiji Fishery Development Limited Company, Sanya 572000 ;
3. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306
黄鳍金枪鱼(Thunnus albacares),隶属鲈形目(Perciformes),鲭亚目(Scombroidei),鲭科(Scombridae),金枪鱼属(Thunnus)。分布于印度洋、太平洋和大西洋的热带、亚热带以及温带广阔海域,在我国主要分布于东海和南海海域。黄鳍金枪鱼具有跨洋洄游性,成群活动的特点,是以鱼类、头足类、甲壳类等为食的大型肉食性鱼类。是金枪鱼类中较为常见、渔获数量较多的一个种类,常在深海区域活动并且游动速度快、活动能力强(林德芳等, 2001),不易受近海环境污染。其鱼肉质鲜美,口感香滑,富含蛋白质、多不饱和脂肪酸和多种维生素,具有较高的营养价值,是世界上名贵的海洋经济鱼类之一(宋利明等, 2004;李娟等, 2008)。黄鳍金枪鱼的产量以捕捞为主,2003年黄鳍金枪鱼占全球金枪鱼捕捞量的62.1%(杨宝瑞, 2007)。目前,黄鳍金枪鱼被世界自然保护联盟(IUCN)列入“接近受威胁”的类别中,资源遭受过度开发。据报道,早在2008年,印度洋、太平洋和大西洋三大洋区的黄鳍金枪鱼资源就处于完全开发状态(黄斌, 2008)。在我国,东海和南海海域也有金枪鱼资源分布,许多远洋渔船已经开始了金枪鱼围网渔业,而我国金枪鱼养殖却处于空白状态(宋利明等, 2004)。
鱼类形态学的性状研究为其生物学分类提供重要依据,并且与种质、行为和食性等紧密相关(王波等, 2002)。鱼类生长有其特定的生长模式,研究其生
长参数和规律,是对鱼类生长性能进行遗传评估的重要指标(李思发,1998),同时也为鱼类养殖提供理论基础。国内外学者对黄鳍金枪鱼的研究多集中在不同海域的资源状况和渔业生物学方面。国外学者对其种群结构(Appleyard et al, 2001; Ward et al, 1997)、分布与海洋环境关系(Brill et al, 1999; Lu et al, 2011)、资源状况(Langley et al, 2009、2011)、生物学特性(Itano, 2000; Lehodey et al, 1999; Sun et al, 2005)等方面进行了研究。宋利明等(2004)对大西洋中部黄鳍金枪鱼的性腺成熟度、摄食等级、摄食种类、性比、叉长与体重关系等生物学特性进行了分析。朱国平等(2006)对印度洋中西部黄鳍金枪鱼不同时期的体长与体重关系、雌雄性比、摄食等级等生物学特性也做了相关研究。由于研究海域及时间不同,结果也不尽相同。对黄鳍金枪鱼的形态特征和生长参数的研究,特别对其幼鱼的生物学指标和特性的分析研究目前尚未见报道。本研究以中国南海三沙市美济礁海域捕获的野生黄鳍金枪鱼幼鱼作为研究对象,测量与其外部形态和生长相关的参数,对其形态特征、生长模式和规律进行分析,为黄鳍金枪鱼种质资源的保护和利用,为我国金枪鱼养殖模式和苗种繁育技术研究,提供理论依据和基础资料。
1 材料与方法 1.1 材料黄鳍金枪鱼于2013年9月取自中国南海美济礁海域(9°55’N,115°32’E)。采用光诱单网捕捞船捕获的黄鳍金枪鱼当龄鱼52尾,体重为195.12-327.21 g,体长为21.0-24.4 cm。
1.2 测量方法采用Kerstan(1995)的测量方法,用游标卡尺和直尺(±0.1cm)对全长、体长、头长、体高、头高、眼径、眼间距、口裂、吻长、尾柄长和尾柄高11个指标进行测量,用电子天平(±0.1 g)称重。记录第1背鳍、第2背鳍、胸鳍、腹鳍、臀鳍、尾鳍和脊椎骨7项数量性状。
1.3 数据分析用Excel 2013和SPSS 19软件进行分析,数据以平均值±标准差(Mean±SD)表示。用标准差/平均值×100%计算变异系数;体重与各形态参数的关系分别以体重为因变量,各参数为自变量进行相关分析,并进行通径分析和多元回归分析。P < 0.01为差异极显著,P < 0.05为差异显著。
计算平均值(x)、标准差(SD)和变异系数(CV),相关系数(rxy),通径系数(Pi),单个自变量决定系数(di)和共同决定系数(dij)。计算公式为:
| $ \begin{array}{*{20}{c}} {\overline x = \frac{{\sum x }}{n};SD = \sqrt {\frac{{\sum {\left({x- \overline x } \right)} }}{{n- 1}}} ;CV\left(\% \right) = \frac{S}{{\overline x }} \times 100\% ;}\\ {{r_{xy}} = \frac{{\sum\limits_{i = 1}^n {\left[{\left({x-\overline x } \right) \times \left({y-\overline y } \right)} \right]} }}{{\sqrt {\sum\limits_{i = 1}^n {{{\left({x -\overline x } \right)}^2} \times \sum\limits_{i = 1}^n {{{\left({y -\overline y } \right)}^2}} } } }};{r_{{x_i}y}} = {P_i} + \sum {{r_{ij}} \times {P_j};} }\\ {{d_i} = P_i^2;{d_{ij}} = 2{r_{ij}} \times P \times {P_j}} \end{array} $ |
式中,x、y为性状测量值,x、y为性状测量平均值,rxy为相关系数,n为测量尾数(王新安等, 2008;耿绪云等, 2011)。
2 结果 2.1 外部形态特征本次采集的样本特征:鱼体呈纺锤形,体形较长、粗壮而圆,横断面略呈圆形,向后逐渐细尖,尾柄细长,尾鳍呈叉状(图 1)。头长略大于体高;眼小,侧上位,眼间隔大于眼径;口大,口裂向下倾斜。体最高处位于第1背鳍基中部,第2背鳍高于第1背鳍。第1背鳍鳍条数为14,第2背鳍鳍条数为14-15,胸鳍鳍条为33-34,腹鳍鳍条为5,臀鳍鳍条为14-15,尾鳍鳍条为28-30;背鳍、臀鳍后方各有一排鳍条数为8的小鳍,每个小鳍上又有8根细小分支。背部较暗,呈深蓝色,腹部银白色,体表具有浅银灰色纵向间隔的条纹,并有彩色明亮的光泽,尾鳍末端呈白色。椎骨为37。全长为体长的1.2倍,体长为体高的3.7倍,为头长的3.5倍;头长为吻长的2.9倍,为眼径的4.3倍,为眼间距的2.8倍;尾柄长为尾柄高的5.7倍。可数性状符合黄鳍金枪鱼形态分类特征。

|
图 1 黄鳍金枪鱼幼鱼的外部形态 Figure 1 Morphology of T. albacares |
形态特征的参数值见表l。从表 1可以看出,在黄鳍金枪鱼幼鱼形态性状中体重的变异系数最大,为14.5%。相关分析结果见表 2。从表 2可以看出,参数间具有差异极显著(P<0.01)的性状包括:全长与体长、全长与头长、全长与体高、全长与体重、体长与体重、头长与体重、体高与尾柄长、体高与尾柄高、体高与体重、眼间距与尾柄高、尾柄高与体重;具有差异显著(P<0.05)的性状有全长与口裂、体长与头长、头长与体高、头长与吻长、体高与头高、体高与口裂、眼径与眼间距、眼径与尾柄高、眼间距与体重、口裂与体重。其余性状间差异不显著(P > 0.05)。
|
|
表 1 黄鳍金枪鱼形态特征的统计值 Table 1 Statistical analysis on the morphological characters of T. albacares |
|
|
表 2 黄鳍金枪鱼形态特征的相关系数 Table 2 The correlation coefficient of morphological characters in T. albacares |
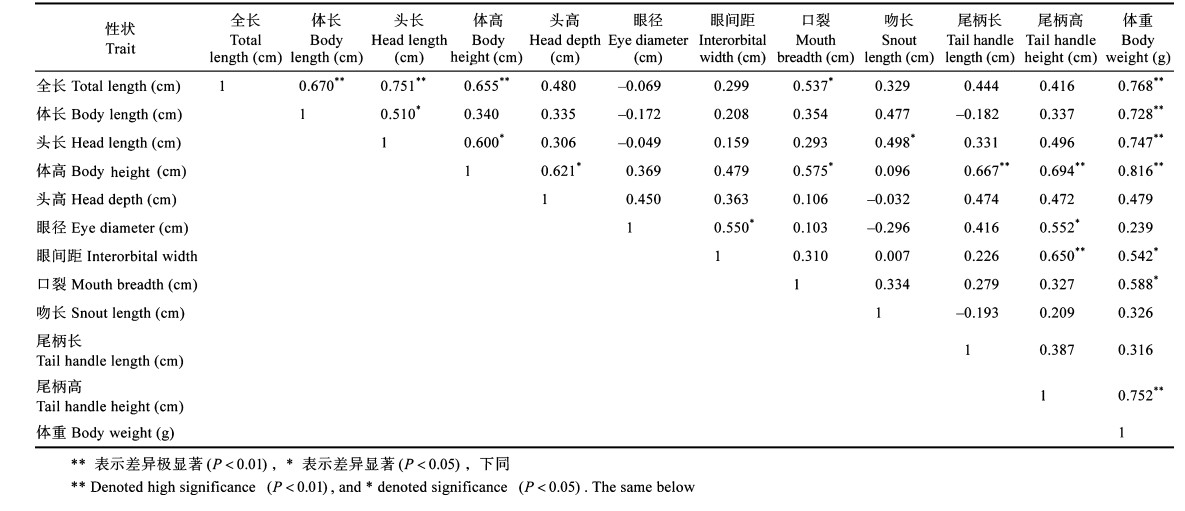
相关关系分析结果显示,与其他参数存在最多相关性是体重、体高,分别与其他7项性状存在显著差异(P < 0.05);存在最少相关性的是头高、吻长、尾柄长,分别与其他1项性状存在显著差异(P < 0.05)。各形态性状与体重相关系数从大到小依次为体高、全长、尾柄高、头长、体长、口裂和眼间距,各性状与体高的相关系数从大到小依次为体重、尾柄高、尾柄长、全长、头高、头长和口裂。
2.3 各性状对体重影响的通径分析在相关关系分析的基础上,将外部形态性状对体重的影响进行通径分析,删除通径系数不显著的性状后,有体长、体高、尾柄高3个性状的通径系数差异极显著,结果见表 3。直接通径系数分别为0.479、0.469、0.266,体长、体高对体重的直接作用大于间接作用的影响,而尾柄高对体重通过体高、体长间接作用大于其直接作用的影响。
|
|
表 3 外部形态性状对体重的通径分析 Table 3 The path coefficients of morphological traits on body weight |
通径分析结果显示,不同性状对体重产生直接作用从大到小依次为体长、体高和尾柄高,对体重产生间接作用从大到小依次为尾柄高、体高和体长。
2.4 各性状对体重的决定系数分析决定系数包括单个性状和各性状间对体重的共同决定系数,在通径分析的基础上对决定系数进行计算,结果见表 4。从表 4可以看出,体长、体高、尾柄高单个性状对体重的决定系数分别为0.229、0.220、0.071,各性状间对体重的决定系数中体高与尾柄高的共同作用最大,为0.173,体长与尾柄高的共同作用最小,为0.086。结果显示,对体重起决定性作用的性状为体长和体高,尾柄高主要通过与体高、体长协同起作用。各形态特征对体重的决定系数总和(∑d)为0.932,说明体长、体高、尾柄高是影响体重的主要性状。
|
|
表 4 外部形态性状对体重的决定系数 Table 4 The determinant coefficients of morphological traits on body weight |
综合2.1-2.4分析,以体重为因变量(y),体高(x1)、体长(x2)和尾柄高(x3)为自变量,对进行多元回归分析,得到估算体重的最优多元回归方程为:
| $ y = 30.482{x_1} + 18.328{x_2} + 199.490{x_3}-500.785 $ |
经显著性检验,体高、体长和尾柄高对体重的偏回归系数及回归常数均达到极显著水平(P < 0.01)。
3 讨论 3.1 影响黄鳍金枪鱼幼鱼体重的主要形态性状分析本研究测得的黄鳍金枪鱼幼鱼形态性状包括全长、体长、头长、体高、头高、眼径、眼间距、口裂、吻长、尾柄长、尾柄高和体重共12项。对其进行相关分析结果显示,存在显著差异的性状对体重的影响大小依次为体高、全长、尾柄高、头长、体长、口裂、眼间距。但由于相关分析不能判断各自变量对因变量所产生影响的大小(Deboski et al, 1999; Hame et al, 2000),因此,在相关分析的基础上,进行通径分析和决定系数分析。通径分析可反映因变量和各自变量的关系,同时,将各自变量的影响效应直接进行比较,并区分各自变量的直接作用和间接作用,可以全面反映各自变量对因变量的相对重要性。结果显示,单个性状和各性状间的共同决定系数之和,即通径系数为0.932,通径分析和决定系数分析结果相一致,∑d>0.85,表明被保留的体长、体高、尾柄高3个性状是影响体重的主要性状,其他被删除的性状对其影响较小,保留的3个性状中,体长、体高主要通过直接作用来对体重产生影响,而尾柄高主要与体高、体长协同对体重起间接作用,以确保体重在生长中起到最具有实际意义的因素。
唐瞻杨等(2010)研究表明,影响尼罗罗非鱼(Oreochromis niloticus)雌鱼体重的主要性状为体长、体高、体宽和头长,影响雄鱼体重的主要性状为体高、体长、躯干长和体宽;影响黄姑鱼(Nibea albiflora) 1龄幼鱼体重的主要形态性状为体长、体高、体厚、尾柄高和眼间距(薛宝贵等, 2011);对于竹荚鱼(Trachurus japonicus)的1龄幼鱼和2龄成鱼,头长、头高、尾柄高、体重这4个参数均与体长存在显著相关关系(胡晓亮等, 2013)。本研究中影响体重的3个主要性状是体长、体高和尾柄高,与其他研究结果较一致。分析表明,这些性状是影响鱼类体重的重点性状,与金枪鱼的体型特征和习性密切相关,尾柄细长,尾部呈半月形,可提供快速游动的动力,使其能够具备快速向前冲刺的能力。
不同生长阶段和性别影响体重的主要性状各不同。唐瞻杨等(2010)研究表明,影响尼罗罗非鱼雌鱼和雄鱼体重的主要性状有所不同;梭鱼(Liza haematocheila)在不同年龄组,影响其体重的重点形态性状明显不同(耿绪云等, 2011);由于美国红鱼(Sciaenops ocellatus)的生长环境、生长阶段等不同,其生长存在阶段性差异,同时不同性别的美国红鱼其体长、体重生长也有差异(王波等, 2002)。因黄鳍金枪鱼较为珍贵并且捕捞难度大,本研究只对黄鳍金枪鱼幼鱼阶段影响体重的主要性状进行分析,对于这些主要性状是否会随生长时期的变化而有不同的影响效果,有待进一步研究。
3.2 多元方程的建立通过测定各形态性状值,发现体重的变异系数最大,说明体重受环境影响产生的系统误差较大,体高、体长、尾柄高的变异系数相对较小,性状较为稳定。以体重为因变量,体高、体长、尾柄高为自变量,进行通径分析和多元分析后建立了估算体重的最优多元回归方程,明确了体重与主要形态性状之间的数量关系,经过回归预测,实际值与估计值之间差异不显著(P>0.05),表明该方程能够较为真实的反映体重与主要性状间的关系,能够应用于实际研究工作中。
| 王波, 刘世禄, 张锡烈, 等. 美国红鱼形态和生长参数的研究. 海洋水产研究 , 2002, 23 (1) : 47-53 | |
| 王新安, 马爱军, 许可, 等. 大菱鲆幼鱼表型形态性状与体重之间的关系. 动物学报 , 2008, 54 (3) : 540-545 | |
| 朱国平, 陈新军, 许柳雄. 印度洋中西部黄鳍金枪鱼生物学特性的初步研究. 海洋渔业 , 2006, 28 (1) : 25-29 | |
| 宋利明, 陈新军, 许柳雄. 大西洋中部金枪鱼延绳钓渔场黄鳍金枪鱼(Thunnus albacares)生物学特性的初步研究. 海洋与湖沼 , 2004, 35 (6) : 538-542 | |
| 李娟, 林德芳, 黄滨, 等. 世界金枪鱼网箱养殖技术现状与展望. 海洋水产研究 , 2008, 29 (6) : 142-147 | |
| 李思发. 中国淡水主要养殖鱼类种质研究. 上海: 上海科学技术出版社, 1998 . | |
| 林德芳, 关长涛, 黄文强. 金枪鱼曳绳拟饵研制与海上试验结果. 海洋水产研究 , 2001, 22 (3) : 42-49 | |
| 杨宝瑞. 日本金枪鱼养殖现状. 中国水产 , 2007 (2) : 25 | |
| 胡晓亮, 陈庆余. 竹荚鱼形态特征参数的比较分析. 现代食品科技 , 2013, 29 (1) : 34-41 | |
| 耿绪云, 马维林, 李相普, 等. 梭鱼(Liza haematocheila)外部形态性状对体重影响效果分析. 海洋与湖沼 , 2011, 42 (4) : 530-537 | |
| 唐瞻杨, 林勇, 陈忠, 等. 尼罗罗非鱼的形态性状对体重影响效果的分析. 大连海洋大学学报 , 2010, 25 (5) : 428-433 | |
| 黄斌. 世界主要金枪鱼类资源状况与管理. 现代渔业信息 , 2008, 23 (1) : 22-25 | |
| 薛宝贵, 辛俭, 楼宝, 等. 黄姑鱼一龄幼鱼形态性状对体重的影响分析. 浙江海洋学院学报(自然科学版) , 2011, 30 (6) : 492-498 | |
| Appleyard SA, Grewe PM, Innes BH, et al. Population structure of yellowfin tuna (Thunnus albacares) in the western Pacific Ocean, inferred from microsatellite loci. Mar Biol , 2001, 139 (2) : 383-393 DOI:10.1007/s002270100578 | |
| Brill RW, Block BA, Boggs CH, et al. Horizontal movements and depth distribution of large adult yellowfin tuna (Thunnus albacares) near the Hawaiian Islands, recorded using ultra-sonic telemetry: implications for the physiological ecology of pelagic fishes. Mar Bio , 1999, 133 (3) : 395-408 DOI:10.1007/s002270050478 | |
| Deboski P, Dobosz S, Robak S, et al. Fat level in body of juvenile Atlantic salmon(Salmo salar L.), and sea trout (Salmo trutta M. trutta L.), and method of estimation from morphometric data. Arch Polish Fish , 1999, 7 (2) : 237-243 | |
| Itano DG. The reproductive biology of yellowfin tuna(Thunnus albacares) in Hawaiian waters and the western tropical Pacific Ocean: project summary. Hawaii: University of Hawaii, Joint Institute for Marine and Atmospheric Research , 2000 : 1-62 | |
| Harue K, Mutsuyshi T, Katsuya M, et al. Estimation of body fat content from standard body length and body weight on cultured Red Sea bream. Fisheries Sci , 2000, 66 (2) : 365-371 DOI:10.1046/j.1444-2906.2000.00056.x | |
| Kerstan M. Age and growth rates of agulhas bank horse mackerel Trachurus capensis-comparison of otolith aging and length frequency analyses. S Afr J Marine Sci , 1995, 15 (1) : 137-156 DOI:10.2989/025776195784156368 | |
| Lu HJ, Lee KT, Lin HL, et al. Spatio-temporal distribution of yellowfin tuna Thunnus albacares and bigeye tuna Thunnus obesus in the Tropical Pacific Ocean in relation to large scale temperature fluctuation during ENSO episodes. Fisheries Sci , 2011, 67 (6) : 1046-1052 | |
| Langley A, Briand K, Kirby DS, et al. Influence of oceanographic variability on recruitment of yellowfin tuna (Thunnus al-bacares) in the western and central Pacific Ocean. Can J Fish Aquatic Sci , 2009, 66 (9) : 1462-1477 DOI:10.1139/F09-096 | |
| Langley A, Hoyle S, Hampton J. Stock assessment of yellowfin tuna in the western and central Pacific Ocean. Pohnpei: Western and Centra1 Pacific Fisheries Commission Scientific Committee , 2011 : 1-33 | |
| Lehodey P, Leroy B. Age and growth of yellowfin tuna (Thunnus albacares) from the western and central Pacific Ocean as indicated by daily growth increments and tagging data. Noumea: Oceanic Fisheries Programme Secretariat of the Pacific Community , 1999 : 2-8 | |
| Sun C, Wang WR, Yeh SZ. Reproductive biology of yellowfin tuna in the Central and Western Pacific Ocean. Noumea: the Scientific Committee of the Western and Centra1 Pacific Fisheries Commission , 2005 : 1-7 | |
| Ward RD, Elliott N, Innes B, et al. Global population structure of yellowfin tuna, Thunnus albacares, inferred from allozyme and mitochondrial DNA variation. Fishery Bulletin , 1997, 95 (3) : 566-575 |