



2. 青岛海洋科学与技术国家实验室 海洋渔业科学与食物产出过程功能实验室 青岛 266071
2. Laboratory for Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071
关键种(Keystone species)是指当群落中单个这样种发生变化,会引起群落结构振荡,甚至导致生态功能紊乱乃至整个系统崩溃,因此,关键种对整个生态系统结构和功能发挥了重要作用。关键种可以是濒危种、土著种、外来种,甚至是广布种(Mills et al, 1993; Libralato et al, 2006; Modlmeiera et al, 2015),并且随着区域、时间、生态系统的变化而不同,环境条件的改变也会导致其更替(Jordán et al, 2002;Gili, 2002),关键种决定了水域生态系统食物网结构变化的方向,了解生态关键种的长期变化有助于解析整个生态系统的演替过程(Pauly et al, 2001)。如何筛选和识别关键种就变得尤为关键,也成为近年来渔业生态学的国际研究热点(Libralato et al, 2006; Jordán, 2009; Coll et al, 2012; Eddy et al, 2014; Modlmeiera et al, 2015; Valls et al, 2015; Torres et al, 2017)。国内对关键种的报道主要是综述性的文章(葛宝明等, 2004; 陈清潮, 2012; 杜洪业等, 2012),有关水域生态系统关键种的筛选仅见于张豫等(2013)、杨涛等(2016)等。本研究基于1959~2015年莱州湾春季渔业资源调查数据,构建了该水域鱼类群落食物网拓扑结构,分析了该水域关键种的长期变化,以期为莱州湾生态系统的演变和渔业资源的适应性管理提供科学依据。
1 材料与方法 1.1 数据来源本研究基于1959年、1982年、1993年、2003年和2015年莱州湾春季(5月)底拖网渔业资源调查数据,其中1959~2003年数据参考金显仕等(1998, 2000, 2001, 2005)、邓景耀等(2000)、Jin(2004)、Jin等(2013)和李显森等(2008)。2015年数据来源于中国水产科学研究院黄海水产研究所973项目莱州湾调查,调查站位延续中国水产科学研究院黄海水产研究所历史调查站位设计方案(图 1),调查船租用205 kW双拖渔船,调查网具参数为网口高度6 m,网口宽度22.6 m,网口周长1740目,网目63 mm,囊网网目20 mm,拖速3 kn,拖网1 h。渔获物中所有鱼类种类均被记录,并留取胃样品进行实验室胃含物分析。食性数据参考邓景耀等(1988a, 1988b)、唐启升等(1990)、金显仕等(1999, 2005)及调查取样的胃含物分析。

|
图 1 莱州湾渔业资源调查站位 Figure 1 Sampling stations in Laizhou Bay |
鱼类群落食物网拓扑结构指标计算参考杨涛等(2016),群落多样性指数计算参照金显仕等(1999),群落种间聚类系数计算参照Watts等(1998)。
鱼类群落食物网拓扑结构节点密度(Density,Dd)、种间关联度(Connectance,C)参照Dunne等(2002),具体如下:
节点密度Dd:
| $ {D_d} = S/L $ |
种间关联度指数C:
| $ C = L/{(S)^2} $ |
式中,S为鱼类群落中种类数量;L为群落内各种间存在的摄食关系的数量,L值越大,说明群落内存在的摄食关系越多;Dd为节点密度,Dd值越大,群落内物种的摄食关系的平均值就越大;C为种间关联度,C值越大,2个物种间存在捕食-被捕食关系的概率就越大。
采用群落离散变量(Fragmentation differences,∆F')和距离权重离散变量(Distance~weighted fragmentation differences,∆DF') (Borgatti, 2013)评估鱼类群落中某些种类的消失对群落结构离散程度的影响,其中,∆ > 0,则群落离散程度增大;∆=0,则离散度不变;∆ < 0,则离散度降低。
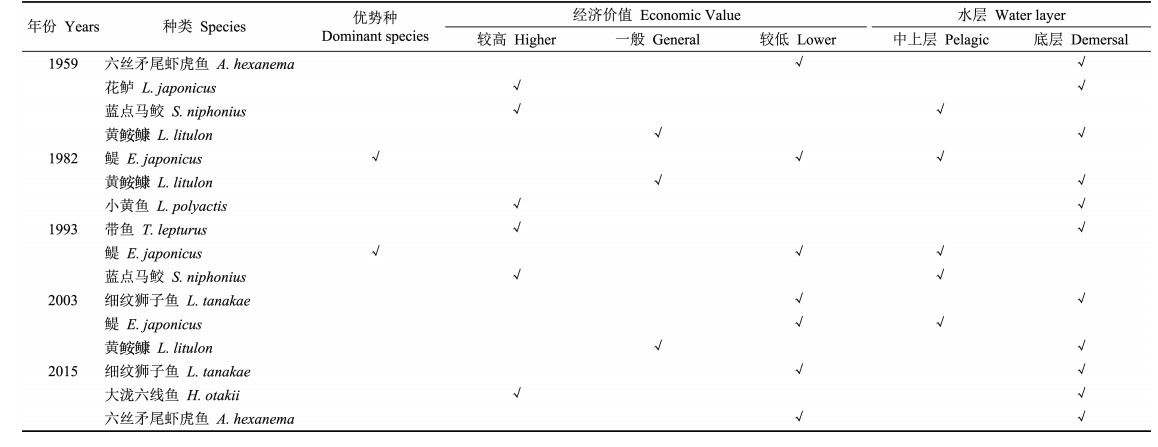
2 结果 2.1 鱼类群落和食物网拓扑结构1959~2015年,莱州湾渔业资源调查共捕获鱼类66种(表 1),隶属于10目34科56属,其中1982年种类最多,共48种,隶属于8目24科42属。其中1982~2015年春季莱州湾鱼类优势种见表 2,1959年优势种为:带鱼、小黄鱼、半滑舌鳎(Cynoglossus semilaevis)、鲬(Platycephalus indicus)和白姑鱼(Pennahia argentata) (金显仕等,2000),可知莱州湾春季鱼类群落优势种群经历了由1950年代的带鱼、半滑舌鳎等重要经济种类向鳀、黄鲫(Setipinna taty)等小型经济种类演替,再经本世纪初的小黄鱼、银鲳(Pampus argenteus)等重要经济物种向六丝矛尾虾虎鱼、短吻红舌鳎(Cynoglossus joyneri)和方氏云鳚(Pholis fangi)等低质底层鱼类演替过程。
|
|
表 1 1959~2015年莱州湾春季鱼类种类组成 Table 1 Fish species composition of the Laizhou Bay in spring during 1959~2015 |
|
|
表 2 1982~2015年莱州湾春季鱼类优势种组成 Table 2 Dominant species composition of the Laizhou Bay in spring during 1982~2015 |
本研究通过丰富度指数(R)、Shannon-Weaver多样性指数(H')和均匀度指数(J')分析了莱州湾鱼类群落的多样性。莱州湾鱼类多样性在年间变化较大(图 2),其中,种类丰富度指数(Rw和Rn分别表示以重量和数量计算的R,H'w、H'n和J'w、J'n与此相同)总体呈下降趋势,1982年最高,随后逐渐降低,至2003年降至最低,然后略有回升。多样性指数总体呈增加趋势,H 'w在1982年后略有降低,2003年后开始回升并超过1982年,而H'n一直升高,但H'n在各年份均未超过H'w。均匀度指数(J 'w和J 'n)在1982到2015年期间变化并不大,但均有少量增加,2015年增至最高。

|
图 2 1959~2015年莱州湾春季鱼类多样性变化 Figure 2 Fish community biodiversity in the Laizhou Bay in spring during 1959~2015 |
1959~2015年,莱州湾春季鱼类食物网包含物种21~46,摄食关系数量70~296个,平均值为146.6,其中,摄食关系数量1982年最多,2003年最低;食物网拓扑结构密度为0.155~0.300,平均密度为0.227,2003年最高,1982年最低;种间关联度为0.140~ 0.182,平均值为0.158,最高值出现在1993年,最低值出现在1982年;聚类系数为0.207~0.326,加权聚类系数为0.194~0.235,平均值分别为0.255和0.214 (图 3)。鱼类食物网拓扑学指标S在1959~2015年期间呈先增加后降低再增加的趋势,但鱼类群落物种数量整体呈下降趋势;L从1959年到1982年呈上升趋势,此后迅速下降,2003年以后保持相对稳定;Dd、C、Cl和W-Cl从1959年到1982年均呈下降趋势,而W-Cl下降相对缓慢,1982年到1993年期间均缓慢增加;1993年Dd和Cl开始上升,而C缓慢下降,W-Cl基本稳定;2003年后Dd、C、Cl和W-Cl均开始下降,Cl降幅最大。这与丰富度指数RW和Rn的变化趋势基本相反,其中Dd与RW呈极显著负相关(P < 0.01),与Rn呈显著负相关(P < 0.05) (表 3)。

|
图 3 1959~2015年莱州湾春季鱼类食物网拓扑结构属性
Figure 3 Topological properties of the fish community in the Laizhou Bay in Spring 1959~2015
L (Links)为摄食关系数量,S (Species)为鱼类群落中物种数量,Dd (Density)为鱼类群落食物网拓扑结构的节点密度,C (Connectance)为鱼类群落中种间关联度,Cl (Clustering coefficient)为聚类系数,W-Cl (Weighted clustering coefficient)为加权聚类系数
L (Links): The number of predator-prey relationships in the food web; S (Species): Species number of fish community; Dd (Density): Node density in food-web topological structure; C (Connectance): Connectance between species in fish community; Cl: Clustering coefficient; W-Cl: Weighted clustering coefficient |
|
|
表 3 莱州湾鱼类多样性指数与食物网拓扑学指标的相关性 Table 3 Correlations between fish biodiversity indices and topological indicators of food web in Laizhou Bay |
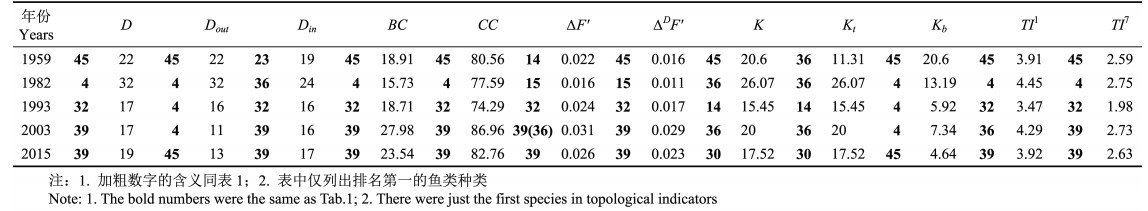
1959年共捕获鱼类34种,其中,短鳍红娘鱼、莱氏舌鳎、星点东方鲀和铅点东方鲀与其他鱼类无任何摄食关系,因此,食物网拓扑结构分析不包括上述4种鱼类。通过构建其余30种鱼类食物网拓扑结构(图 4a),计算其拓扑学指标发现,六丝矛尾虾虎鱼与群落中22种鱼类存在被捕食关系,即D=Dout=22,是该群落中重要的饵料鱼类;花鲈可以捕食群落中其他19种鱼类,即D=Din=19,是该群落的顶级捕食者。同时,六丝矛尾虾虎鱼的BC、TI1、TI7、CC和∆DF'在30种鱼类中最大,说明其对群落内信息交换的控制能力及扩散能力最强、传递信息的速度最快,当六丝矛尾虾虎鱼从该群落剔除时,群落距离权重离散变量增加且增加量最大;但蓝点马鲛从该群落剔除时群落离散变量增加且增加量最大(∆F'=0.022)。在群落中,六丝矛尾虾虎鱼的K和Kb值最高,K=Kb=20.60,表明六丝矛尾虾虎鱼对群落能量流动和信息传递影响最大,其可以通过上行控制效应影响种间作用进而影响整个群落结构。黄鮟鱇的Kt最大,为11.31,说明其对群落下行控制效应最大,即黄鮟鱇通过摄食其他鱼类对整个群落产生调节作用(表 4)。

|
图 4 莱州湾鱼类群落食物网拓扑结构(1959~2015) Figure 4 Topological structure of food-web in the Laizhou Bay fish community during 1959~2015 1) a:1959年;b:1982年;c:1993年;d:2003年;e:2015年;2)图中数字代表鱼类种类,见表 1,下同 1) a: 1959; b: 1982; c: 1993; d: 2003; e: 2015; 2) The numbers represented different kinds of fish species, the details were shown in Tab. 1, the same below |
|
|
表 4 1959~2015年莱州湾春季鱼类群落拓扑结构指标 Table 4 Topological indicators of food web in the Laizhou Bay fish community in spring 1959~2015 |
|
|
表 5 1959~2015年莱州湾春季鱼类群落关键种组成 Table 5 Keystone soecies composition of fish community in Laizhou Bay in spring during 1959~2015 |
1982年共捕获鱼类48种,其中,中颌棱鳀和普氏吻虾虎鱼与该群落其余46种鱼类无任何摄食关系,因此,食物网拓扑结构分析不包括上述2种鱼类。通过构建46种鱼类食物网拓扑结构(图 4b)、计算出其拓扑学指标发现,鳀与群落中32种鱼类存在被捕食关系,即D=Dout=32,是该群落中重要的饵料鱼类;黄鮟鱇Din值最大为24,黄鮟鱇可以摄食群落中其他24种鱼类,D=Din=24,是该群落的顶级捕食者。另外,鳀的BC、TI1、TI7和CC值最大,即46种鱼类中鳀对群落内信息交换的控制能力及扩散能力最强、传递信息速度最快;小黄鱼的∆F'和∆DF'的值最大,分别为0.016和0.01,当小黄鱼从该群落剔除时,该群落结构离散变量和距离权重离散变量均会增加且增加量最大;黄鮟鱇的K和Kt值最大,这表明其对该群落能量流动和信息传递影响最大,且该影响全部来自于下行控制效应(K=Kt=26.07),即黄鮟鱇通过摄食其他鱼类作用于整个群落结构;鳀的Kb值最大,即鳀通过上行控制效应影响着该群落的结构平衡和稳定(表 4)。
1993年共捕获鱼类29种,其中,丝虾虎鱼和尖海龙与该鱼类群落其余27种鱼类无任何摄食关系,因此,不将上述2种鱼类作为研究对象。通过构建其余27种鱼类食物网拓扑结构(图 4c)、计算出其拓扑学指标发现,带鱼与群落中17种鱼类存在捕食和被捕食关系,D=Dout+Din=17,其中,Dout=1,Din=16,这说明带鱼主要以捕食者的角色存在于该鱼类群落,被其捕食的种类达16种,但也会出现被其他鱼类捕食的情况,如被蓝点马鲛捕食。因此,带鱼不仅可以通过捕食其他鱼类影响群落结构,还可以改变自身种群密度影响鱼类群落结构。鳀和长蛇鲻与16种鱼类均存在摄食关系,其中,鳀为被捕食者(D=Dout=16),是该群落中重要的饵料鱼类,而长蛇鲻在该群落中的生态地位与带鱼相似,摄食关系共包括2部分:D= Dout+Din,Dout=15,Din=1。另外,27种鱼类中带鱼的BC、TI1、TI7、CC、∆F'和∆DF'值均最大,即其对群落内信息交换的控制能力及扩散能力最强、传递信息速度最快,当带鱼从该群落中剔除时,群落结构的离散变量和距离权重离散变量均增加,且增加量最大。蓝点马鲛的K和Kt值最大,K=Kt=15.45,蓝点马鲛对该群落能量流动和信息传递影响最大,且该影响全部来自于下行控制效应。鳀的Kb值最大为5.92,说明鳀对该群落的上行控制效应最大,因此,鳀通过生物量的变化影响与其他鱼类的种间关系,进而作用于群落结构的稳定(表 4)。
2003年共捕获鱼类21种,通过构建鱼类食物网拓扑结构(图 4d)、计算出其拓扑学指标发现,细纹狮子鱼与该群落17种鱼类存在摄食关系,D=Dout+Din,其中,Dout=1,Din=16,细纹狮子鱼主要以捕食者的角色存在于该群落,被其捕食的种类达16种,也会出现被其他鱼类捕食的情况,如被黄鮟鱇捕食。黄鮟鱇与群落中15种鱼类存在摄食关系,D=Din=15,是该群落的顶级捕食者,主要依靠捕食其他鱼类影响群落结构的稳定。鳀的Dout值最大为11,D=Dout=11,是该群落中重要的饵料鱼类。群落中细纹狮子鱼的BC、TI7、CC、∆F'和∆DF'值最大,即其对群落内信息交换的控制能力、间接扩散能力最强、信息传递速度最快,当细纹狮子鱼从该群落中剔除时,群落结构的距离权重离散变量增加且增加量最大。黄鮟鱇的∆F'值与细纹狮子鱼相同,当黄鮟鱇和细纹狮子鱼从该群落剔除时,群落离散程度的增加量相同。另外,黄鮟鱇的TI1、K和Kt均最大,K=Kt=20,其对该群落内信息直接传递速度最快,对该群落的能量流动和信息传递影响最大,且该影响全部来自于下行控制效应。鳀的Kb最大,即鳀对该群落的上行控制效应最大(表 4)。
2015年共捕获鱼类28种,其中,银鲳、丝虾虎鱼和裸项栉虾虎鱼与其他25种鱼类无摄食关系,因此,不将上述3种鱼类作为研究对象。通过构建其余25种鱼类群落的食物网拓扑结构(图 4e)、计算其拓扑学指标发现,细纹狮子鱼与群落中19种鱼类存在摄食关系,D=Dout+Din,Dout=2,Din=17,是主要的捕食者,也会被黄鮟鱇和大泷六线鱼捕食。六丝矛尾虾虎鱼的Dout值最大为13,D=Dout=13,即群落中13种鱼类可以捕食六丝矛尾虾虎鱼,六丝矛尾虾虎鱼是该群落中重要的饵料鱼类。25种鱼类中细纹狮子鱼的BC、TI1、TI7、CC、∆F'和∆DF'值最大,其对群落内信息交换的控制能力及扩散能力最强、信息传递速度最快,当细纹狮子鱼从该群落剔除时,群落结构的离散变量和距离权重离散变量均增加且增加量最大。大泷六线鱼对该群落的能量流动和信息传递影响最大,且该影响全部来自于下行控制效应(K=Kt=17.52)。六丝矛尾虾虎鱼的Kb=4.64最大,其对群落的上行控制效应最大(表 4)。
综合上述分析结果,取各指标中最大种类为关键种(Jordán, 2006),1959~2015年莱州湾春季鱼类群落关键种见表 5。
3 讨论近60年来,莱州湾水域春季鱼类群落结构发生了明显改变,鱼类种群趋于小型化、低质化,群落结构组成趋于简单化,平均营养级明显下降(Shan et al, 2013; 张波等, 2015)。生物多样性变化较大,种类丰度指数整体呈下降趋势,多样性指数和均匀度指数整体呈增加趋势。生态群落结构的改变会对鱼类群落食物网的结构和功能产生一定影响,如人类活动对海洋生物资源的选择性开发已导致种群结构与数量的变动,使得全球海洋渔获物的营养级由50年代的3.3下降至1994年的3.1 (Pauly et al, 1998)。本研究中,Dd与RW呈极显著负相关(P < 0.01),与Rn呈显著负相关(P < 0.05),一定程度上可以反映出鱼类群落多样性的变化直接影响着群落食物网的结构和功能。而食物网结构可以帮助确定生态系统中物种的营养关系,并能够评估在不同营养级种类间直接或间接影响(Jordán, 2001; Kitchell et al, 2002; Navia et al, 2010; Bornatowski et al, 2014))。莱州湾鱼类群落食物网经历了以鱼食性种类为主的食物网到以底栖生物食性种类为主的食物网的演变过程(张波等, 2015)。1959~ 2015年莱州湾春季鱼类群落食物网拓扑结构中Dd的取值为0.155~0.300,最高值出现在2003年,最低值出现在1982年,不同年间存在较大波动情况,这与巴西南部巴拉那沿岸等其他的海洋生态系统的食物网拓扑结构相似(Dunne et al, 2002; Gaichas et al, 2008; Bornatowski et al, 2014);而在Cohen等(1984)早期研究结果认为,物种多样性不同的食物网可能具有相同的种间关联度,但因该研究的数据质量和研究方法等原因使得这一研究结果受到Riede等(2010)的质疑。在本研究中莱州湾春季鱼类群落的关联度值为0.140~0.182,较为稳定,在0.03~0.30的范围以内,基本可以排除该群落内种群的摄食可能存在某种特定关系或群落受到外界因素干扰等情况(Dunne et al, 2002; Bornatowski et al, 2014);而过于极端取值可能出现上述情况,如Martinez等(1999)在对Grassland生态群落摄食关系的研究中,C值相对较小为0.026,其原因可能是该生态群落中作为被捕食者的基层物种较少,群落能量流动的来源过于单一;而在Polis(1991)对Coachella Valley的研究中,因受人为因素影响导致C值相对偏大。因此,本研究认为1959~ 2016年莱州湾春季鱼类群落符合自然条件下的群落种间摄食关系。
海洋中的物质能量通过食物链(食物网)转化为海洋生态系统各营养层次的生产力,同时,海洋生态系统对各种物理化学过程的响应通常表现为食物网结构的变化。关键种的生物量未必较高,并不依靠生物量影响群落结构,而是通过种群间的摄食关系密切联系群落的其他种群,并经过食物网的拓扑关系控制着群落的结构和能流,而优势种最明显的特征则是依靠庞大的生物量影响群落结构(Power et al, 1996)。本研究中发现莱州湾5个不同时期的关键种中,既是关键种又是优势种的仅有鳀1个种类,分别为1982年、1993年和2003年莱州湾鱼类群落关键种。而1982年、1993年鳀既是关键种又是优势种,2003年鳀是关键种但非优势种。关键种在一定时间内对其他物种的分布起着直接或间接的调控作用,其存在对于维持群落的种类组成、生态群落的功能和物种多样性等比其他的物种更重要,对生物群落稳定性、物种多样性和许多生态过程的持续和改变起决定性作用,是生态系统的物质循环、能量流动和生物量的调控者。关键捕食者可以通过捕食作用控制着群落中其他重要食物竞争者和捕食者密度,关键被捕食者则可以通过维持捕食者的密度来限制另外被捕食者的密度,是鱼类群落种间关系的第一纽带,其种群数量的变动影响着其他种群的生长、发育和繁殖,是决定鱼类群落动态的重要因素(陈大刚, 1997)。关键捕食者的能量流动与转换又是影响该水域生态系统结构与功能稳定的重要因素,其演变趋势对鱼类群落结构组的发展有一定的诱导作用。
过度捕捞导致近海渔业资源严重衰退,鱼群抵御环境变化的能力降低,近海资源问题日益凸显(王跃中等, 2012; 金显仕等, 2015; 张波等, 2015)。Pitcher (2001)认为受人类活动的影响,20世纪60年代以来须鲸类濒临灭绝,而人类早期的捕捞活动可能已经导致水域生态中关键种濒临灭绝。而过度捕捞将会导致食物网中的关键种由海洋哺乳类、鳕鱼类、鮟鱇鱼类以及大型中上层鱼类向小型底栖无脊椎动物、小型中上层鱼类转变(Coll et al, 2009),本研究中莱州湾春季鱼类群落关键种由经济价值较高的花鲈、小黄鱼和蓝点马鲛等演变为以细纹狮子鱼、鳀和六丝矛尾虾虎鱼等经济价值较低的种类;同时,关键种的栖息环境也由中上层与底层生境(蓝点马鲛、花鲈、鳀等)演变为底层单一生境(细纹狮子鱼和六丝矛尾虾虎鱼);关键种的这种更替导致食物网拓扑结构向简单化发展,在某种程度上也增加了鱼类群落结构脆弱性。气候变化直接影响渔业生物的多样性和分布,而全球变暖将通过对近岸湿地生境的破坏,进而破坏众多鱼类的育肥场所,而水温变化将直接影响鱼类的生长、摄食、产卵及洄游等,驱使在河口或沿岸产卵的鱼类向较冷的地区洄游(樊伟等, 2001; 王跃中等, 2012; 单秀娟等, 2017),是影响鱼类群落结构变化的重要因素,也是改变的群落关键种的潜在因素。Drezner (2013)认为一定条件下水源是限制陆地生态系统中关键物种仙人掌(Carnegiea gigantea)分布的重要环境因素。而水域生态系统中,Sanford (1999)的研究表明野外和室内实验中水温降低,关键捕食者赭色海星(Pisaster ochraceus)对主要被捕食者的捕食明显减弱。因此,在某一生态群落中关键种受多种外界因素的影响,如环境变化、人为活动等,由于物种洄游性季节分布导致鱼类群落结构的时序相对不稳定,因此季节变化是影响关键种的不可忽视的重要因素,关键种存在季节性分布的可能,后续继续深入对鱼类群落结构关键种季节性变化的研究。
| Borgatti SP. Analyzing social networks. Sage, 2013 | |
| Bornatowski H, Navia AF, Braga RR, et al. Ecological importance of sharks and rays in a structural foodweb analysis in southern Brazil. ICES Journal of Marine Science, 2014, 71(7): 1586-1592 DOI:10.1093/icesjms/fsu025 | |
| Chen DG. Fishery resources biology. Beijing: China Agricultrue Press, 1997: 80-100. | |
| 陈大刚. 渔业资源生物学. 北京: 中国农业出版社, 1997: 80-100. | |
| Chen QC. Conservation and sustainable development of marine keystone species. Green Leaf, 2012(10): 35-38 [陈清潮. 保护海洋关键物种与可持续发展. 绿叶, 2012(10): 35-38] | |
| Cohen JE, Briand F. Trophic links of community food webs. Proceedings of the National Academy of Sciences of the United States of America, 1984, 81(13): 4105-4109 DOI:10.1073/pnas.81.13.4105 | |
| Coll M, Libralato S. Contributions of food web modelling to the ecosystem approach to marine resource management in the Mediterranean Sea. Fish and Fisheries, 2012, 13(1): 60-88 | |
| Coll M, Palomera I, Tudela S. Decadal changes in a NW Mediterranean Sea food web in relation to fishing exploitation. Ecological Modelling, 2009, 220(17): 2088-2102 DOI:10.1016/j.ecolmodel.2009.04.049 | |
| Deng JY, Jin XS. Study on fishery biodiversity and its conservation in Laizhou Bay and Yellow River Estuary. Zoological Research, 2000, 21(1): 76-82 [邓景耀, 金显仕. 莱州湾及黄河口水域渔业生物多样性及其保护研究. 动物学研究, 2000, 21(1): 76-82] | |
| Deng JY, Meng TX, Ren SM, et al. Species composition, abundance and distribution of fishes in the Bohai Sea. Marine Fisheries Research, 1988a, 9: 10-89 [邓景耀, 孟田湘, 任胜民, 等. 渤海鱼类种类组成及数量分布. 海洋水产研究, 1988a, 9: 10-89] | |
| Deng JY, Meng TX, Ren SM. Food web of fishes in the Bohai Sea. Marine Fisheries Research, 1988b, 9: 151-171 [邓景耀, 孟田湘, 任胜民. 渤海鱼类的食物关系. 海洋水产研究, 1988b, 9: 151-171] | |
| Drezner TD. The paradoxical distribution of a shallow~rooted keystone species away from surface water, near the water~ limited edge of its range in the Sonoran Desert: Seed~ seedling conflicts. Acta Oecologica, 2013, 47: 81-84 DOI:10.1016/j.actao.2013.01.003 | |
| Du HY, Xu CY. Research progress of the relationship between keystone species and the dynamic of community in forestry ecosystem. World Forestry Research, 2012, 25(5): 35-39 [杜洪业, 徐程扬. 森林生态系统中关键种与群落动态的关系研究进展. 世界林业研究, 2012, 25(5): 35-39] | |
| Dunne JA, Williams RJ, Martinez ND. Food~web structure and network theory: the role of connectance and size. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99: 12917-12922 DOI:10.1073/pnas.192407699 | |
| Eddy TD, Pande A, Gardner JPA. Massive differential site~ specific and species~specific responses of temperate reef fishes to marine reserve protection. Global Ecology and Conservation, 2014, 1: 13-26 | |
| Fan W, Cheng YH, Shen XQ. Effects of global environment change and human activity on fishery resources. Journal of Fishery Sciences of China, 2001, 8(4): 91-94 [樊伟, 程炎宏, 沈新强. 全球环境变化与人类活动对渔业资源的影响. 中国水产科学, 2001, 8(4): 91-94] | |
| Gaichas SK, Francis RC. Network models for ecosystem-based fishery analysis: a review of concepts and application to the Gulf of Alaska marine food web. Canadian Journal of Fisheries and Aquatic Sciences, 2008, 65: 1965-1982 DOI:10.1139/F08-104 | |
| Ge BM, Bao YX, Zheng X. A review on key species study in ecology. Chinese Journal of Ecology, 2004, 23(6): 102-106 [葛宝明, 鲍毅新, 郑祥. 生态学中关键种研究综述. 生态学杂志, 2004, 23(6): 102-106] | |
| Gili JM. Towards a transitory or ephemeral key habitat concept. Trends in Ecology and Evolution, 2002, 17(10): 453 | |
| Jin XS, Deng JY. Variations in community structure of fishery resources and biodiversity in the Laizhou Bay. Chinese Biodiversity, 2000, 8(1): 65-72 [金显仕, 邓景耀. 莱州湾渔业资源群落结构和生物多样性的变化. 生物多样性, 2000, 8(1): 65-72] | |
| Jin XS, Deng JY. Yearly variations of fishery resources and biodiversity in the Laizhou Bay. Marine Fisheries Research, 1999, 20(1): 6-12 [金显仕, 邓景耀. 莱州湾春季渔业资源及生物多样性的年间变化. 海洋水产研究, 1999, 20(1): 6-12] | |
| Jin XS, Dou SZ, Shan XJ, et al. Hot spots of frontiers in the research of sustainable yield of chinese inshore fishery. Progress in Fishery Sciences, 2015, 36(1): 124-131 DOI:10.11758/yykxjz.20150119 [金显仕, 窦硕增, 单秀娟, 等. 我国近海渔业资源可持续产出基础研究的热点问题. 渔业科学进展, 2015, 36(1): 124-131] | |
| Jin XS, Shan XJ, Li XS, et al. Long-term changes in the fishery ecosystem structure of Laizhou Bay, China. Science China Earth Sciences, 2013, 56(3): 366-374 DOI:10.1007/s11430-012-4528-7 | |
| Jin XS, Tang QS. The structure, distribution and variation of the fishery resources in the Bohai Sea. Journal of Fishery Sciences of China, 1998, 5(3): 18-24 [金显仕, 唐启升. 渤海渔业资源结构、数量分布及其变化. 中国水产科学, 1998, 5(3): 18-24] | |
| Jin XS, Zhao XY, Meng TX, et al. The yellow sea and bohai sea biological resources and habitats. Beijing: Science Press, 2005: 262-351. [金显仕, 赵宪勇, 孟田湘, 等. 黄、渤海生物资源与栖息环境. 北京: 科学出版社, 2005: 262-351.] | |
| Jin XS. Long-term changes in fish community structure in the Bohai Sea, China. Estuarine, Coastal and Shelf Science, 2004, 59(1): 163-171 DOI:10.1016/j.ecss.2003.08.005 | |
| Jin XS. The dynamics of major fishery resources in the Bohai Sea. Journal of Fishery Sciences of China, 2001, 7(4): 22-26 [金显仕. 渤海主要渔业生物资源变动的研究. 中国水产科学, 2001, 7(4): 22-26] | |
| Jordán F, Liu WC, Davis AJ. Topological keystone species: measures of positional importance in food webs. Oikos, 2006, 112(3): 535-546 DOI:10.1111/oik.2006.112.issue-3 | |
| Jordán F, Scheuring I. Searching for keystones in ecological networks. OIKOS, 2002, 99(3): 607-612 DOI:10.1034/j.1600-0706.2002.11889.x | |
| Jordán F. Keystone species and food webs. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 2009, 364(1524): 1733-1741 DOI:10.1098/rstb.2008.0335 | |
| Jordán F. Trophic fields. Community ecology, 2001, 2: 181-185 DOI:10.1556/ComEc.2.2001.2.5 | |
| Kitchell JF, Essington TE, Boggs CH, et al. The role of sharks and longline fisheries in a pelagic ecosystem of the Central Pacific. Ecosystems, 2002, 5: 202-216 DOI:10.1007/s10021-001-0065-5 | |
| Li XS, Niu MX, Dai FQ. The characteristics of structure and distribution of spawning stock of fishery species in the Bohai Sea. Marine Fisheries Research, 2008, 29(4): 15-21 [李显森, 牛明香, 戴芳群. 渤海渔业生物生殖群体结构及其分布特征. 海洋水产研究, 2008, 29(4): 15-21] | |
| Libralato S, Christensen V, Pauly D. A method for identifying Keystone species in food web models. Ecological Modelling, 2006, 195(3-4): 153-171 DOI:10.1016/j.ecolmodel.2005.11.029 | |
| Martinez ND, Hawkins BA, Dawah HA, et al. Effects of sampling effort on characterization of food web structure. Ecology, 1999, 80: 1044-1055 | |
| Mills LS, Soule ME, Doak DF. The keystone-species concept in ecology and conservation. BioScience, 1993, 43(4): 219-224 DOI:10.2307/1312122 | |
| Modlmeiera AP, Laskowskib KL, Brittinghama HA, et al. Adult presence augments juvenile collective foraging in social spiders. Animal Behaviour, 2015, 109: 9-14 DOI:10.1016/j.anbehav.2015.07.033 | |
| Navia AF, Cortés E, Mejía-Falla PA. Topological analysis of the ecological importance of elasmobranch fishes: a food web study on the Gulf of Tortugas, Colombia. Ecological Modelling, 2010, 221: 2918-2926 DOI:10.1016/j.ecolmodel.2010.09.006 | |
| Pauly D, Christensen VV, Dalsgaard J, et al. Fishing down marine food webs. Science, 1998, 279(5352): 860-863 DOI:10.1126/science.279.5352.860 | |
| Pauly D, Palomares ML, Froese R, et al. Fishing down Canadian aquatic food webs. Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(1): 51-62 DOI:10.1139/f00-193 | |
| Pitcher TJ. Fisheries managed to rebuild ecosystems? Reconstructing the past to salvage the future. Ecological Applications, 2001, 11(2): 601-617 | |
| Polis GA. Complex desert food webs: An empirical critique of food web theory. American Naturalist, 1991, 138: 123-155 DOI:10.1086/285208 | |
| Power ME, Tilman D, Estes JA, et al. Challenges in the quest for keystones. BioScience, 1996, 46(8): 609-620 DOI:10.2307/1312990 | |
| Riede JO, Rall BC, Banasek-Richter C, et al. Scaling of food-web properties with diversity and complexity across ecosystems. In: Woodward G (ed): Advances in ecological research. Burlington: Academic Press, 2010, 139-170 | |
| Sanford E. Regulation of keystone predation by small changes in ocean temperature. Science, 1999, 283(5410): 2095-2097 DOI:10.1126/science.283.5410.2095 | |
| Shan XJ, Chen YL, Jin XS. Projecting fishery ecosystem health under climate change scenarios: Yangtze River Estuary and Yellow River Estuary. Progress in Fishery Sciences, 2017, 38(2): 1-7 DOI:10.11758/yykxjz.20160407001 [单秀娟, 陈云龙, 金显仕. 气候变化对长江口和黄河口渔业生态系统健康的潜在影响. 渔业科学进展, 2017, 38(2): 1-7] | |
| Shan XJ, Sun PF, Jin XS, et al. Long-term changes in fish assemblage structure in the Yellow River Estuary ecosystem, China. Marine and Coastal Fisheries: Dynamics, Management and Ecosystem Science, 2013, 5: 65-78 DOI:10.1080/19425120.2013.768571 | |
| Tang QS, Ye MZ. The exploitation and conservation of nearshore fisheries resources off Shandong. Beijing: China Agriculture Press, 1990: 90-115. [唐启升, 叶懋中. 山东近海渔业资源开发与保护. 北京: 农业出版社, 1990: 90-115.] | |
| Torres MA, Casini M, Huss M, et al. Food-web indicators accounting for species interactions respond to multiple pressures. Ecological Indicators, 2017, 77: 67-69 DOI:10.1016/j.ecolind.2017.01.030 | |
| Valls A, Coll M, Christensen V. Keystone species: toward an operational concept for marine biodiversity conservation. Ecological Monographs, 2015, 85(1): 29-47 DOI:10.1890/14-0306.1 | |
| Wang YZ, Sun DR, Lin ZJ, et al. Analysis on responses of hairtail catches to fishing and climate factors in the Yellow Sea and Bohai Sea, China. Journal of Fishery Sciences of China, 2012, 19(6): 1043-1050 [王跃中, 孙典荣, 林昭进, 等. 捕捞压力和气候因素对黄渤海带鱼渔获量变化的影响. 中国水产科学, 2012, 19(6): 1043-1050] | |
| Watts DJ, Strogatz SH. Collective dynamics of 'small-world' networks. Nature, 1998, 393(6684): 440-442 DOI:10.1038/30918 | |
| Yang T, Shan XJ, Jin XS, et al. The keystone species of fish community in the Laizhou Bay. Journal of Fisheries of China, 2016, 40(10): 1613-1623 [杨涛, 单秀娟, 金显仕, 等. 莱州湾鱼类群落关键种. 水产学报, 2016, 40(10): 1613-1623] | |
| Zhang B, Wu Q, Jin XS. Interannual variation in the food web of commercially harvested species in Laizhou Bay from 1959 to 2011. Journal of Fishery Sciences of China, 2015, 22(2): 278-287 [张波, 吴强, 金显仕. 1959~2011年莱州湾渔业资源群落食物网结构的变化. 中国水产科学, 2015, 22(2): 278-287] | |
| Zhang Y, Guo FQ, Hu SJ, et al. Priority of conservation of keystone species of aquatic ecosystem in Huizhou reach of the Dongjiang main river. Journal of Anhui University (Natural Science Edition), 2013, 37(5): 100-108 [张豫, 郭凤清, 胡顺军, 等. 东江干流(惠州段)水生生态系统关键物种的优先保护顺序. 安徽大学学报(自然科学版), 2013, 37(5): 100-108] |