



2. 上海海洋大学 水产种质资源发掘与利用教育部重点实验室 水产科学国家级实验教学示范中心海洋动物系统分类与进化上海高校重点实验室 上海 201306;
3. 广东省渔业种质保护中心 广州 511453
2. Shanghai Ocean University, Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources, Ministry of Education; National Demonstration Center for Experimental Fisheries, Science Education; Key Laboratory of Marine Animal Taxonomy and Evolution, Shanghai 201306;
3. Guangdong Provincial Fishery Germplasm Conservation Center, Guangzhou 511453
盐度是影响海水硬骨鱼类生存的重要环境因子之一。不同鱼类对环境盐度有特定的适应范围。为应对外界盐度的改变,大多数硬骨鱼类进化形成了完备的渗透压调节功能。广盐性鱼类能在较大盐度范围下生活,在盐度发生变化时,相关组织结构会发生适应性变化,从而保持较好的摄食、吸收和能量转化效率。鳃是鱼类的主要渗透调节器官,参与调节机体渗透压、pH、离子浓度、耗氧率及氨氮排泄等,以维持机体内外环境平衡(Shivakumar et al, 1986; Ern et al, 2018)。国内外学者已在红姑鱼(Sciaenops ocellatus)、鲤鱼(Cyprinus carpio)、斑尾复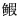
花鲈(Lateolabrax maculatus)隶属于鲈形目(Perciformes)、
实验用鱼采自中国水产科学研究院南海水产研究所珠海基地,为淡水培育幼苗,暂养于蓝色养殖箱中(55 cm×30 cm×42 cm),养殖用水为曝气处理的自来水,盐度为0,水温为27℃~29℃,连续24 h充气,每日定时投喂2次配合饲料,日换水量为总水量的1/3,暂养时间为7 d。暂养结束后,挑选体格健壮,规格相近[平均体长为(4.01±0.27) cm]的个体进行实验。
1.2 实验设计实验设5个实验组S0、S10、S15、S20和S30,对应盐度分别为0、10、15、20和30,其中,S0为对照组,每组设3个平行,每个实验组放幼鱼50尾。实验用水为曝气自来水与净化处理的自然海水调配而成。实验开始前,盐度每隔24 h提高5,用盐度计进行校准,误差不超过0.5,达到相应盐度后开始实验。实验期间,充气、水温控制、换水量及投喂管理与暂养时相同。实验时间为30 d。
1.3 实验方法实验结束后,在各实验组随机取样,取幼鱼右侧鳃丝、脾脏及躯干部右侧肌肉组织样品,于4%多聚甲醛溶液中固定24 h。将固定好的组织样品制成石蜡切片,常规HE染色,中性树脂封片,Zeiss显微镜下观察并拍照。
1.4 数据测量及分析采用Image-ProPlus 5.1软件测量鳃丝宽度、鳃小片长度、鳃小片宽度、鳃小片间距;参考秦桂香等(2010)的方法测量肌纤维的长径和短径,以实验用显微镜(200×)拍摄的图片(视野的一个内切矩形)为单位面积计算肌纤维密度。测量所得数据均用SPSS 22.0软件和Excel 2010软件进行统计学分析。
2 结果 2.1 不同盐度条件下花鲈幼鱼存活率不同盐度条件下花鲈幼鱼养殖死亡率见表 1,除盐度为30条件下有3尾鱼死亡外,其他组均无死亡。
|
|
表 1 不同盐度下花鲈幼鱼的死亡率 Tab.1 Mortality of L. maculatus juveniles under different salinities |
盐度为0时,花鲈幼鱼鳃丝排列紧密,形状较长,顶端膨大呈棒状,内有大量的血细胞;鳃丝两侧有较长的扁平囊状鳃小片平行排列;鳃小片由扁平细胞、柱细胞、血细胞及泌氯细胞组成,细胞饱满,排列紧密;有少许泌氯细胞分布于鳃小片基部,近乎椭圆形,体积较小细胞核呈蓝紫色,细胞质着色较浅;鳃小片上有较多的血细胞,扁平细胞较厚,排列紧密(图 1.1)。盐度为10时,泌氯细胞增大,数量增多,血细胞减少(图 1.2);鳃丝宽度显著小于S0组,鳃小片变宽,扁平细胞增厚,其他结构无明显变化(表 2)。盐度为15时,鳃丝较长,排列整齐,宽度显著变窄(P < 0.05);鳃小片间距增大,长度缩短,宽度变窄,与S0、S10组均差异显著(P < 0.05);泌氯细胞体积增大,数量增多(图 1.3)。盐度为2时,鳃丝较S15组宽,鳃小片减少,鳃小片间距较宽,长度缩短(表 2);泌氯细胞体积明显增大,细胞排列疏松,扁平上皮细胞有脱落溶解现象(图 1.4)。盐度为30时,鳃丝宽度较大,出现断裂脱落现象,鳃丝上细胞排列疏松,泌氯细胞明显膨大,见溶解状态(图 1.5);鳃小片数量较少,形态不规则,小片间距和长度显著增大(P < 0.05) (表 1)。

|
图 1 盐度对花鲈幼鱼鳃的影响
Fig.1 The effect of salinity on the gills of L. maculatus juveniles
1:盐度0;2:盐度10;3:盐度15;4:盐度20;5:盐度30 BC:血细胞;CSC:泌氯细胞;GL:鳃小片;PVC:扁平上皮细胞;PC:柱细胞;CC:软骨细胞 1: Salinity 0; 2: Salinity 10; 3: Salinity 15; 4: Salinity 20; 5: Salinity 30 BC: Blood cell; CSC: Chloride secreting cell; GL: Gill lamellae; PVC: Pavement cell; PC: Pillar cell; CC: Chondrocyte |
|
|
表 2 盐度对花鲈幼鱼鳃组织测量性状的影响 Tab.2 The effect of salinity on the gill meristic characters of L. maculatus juveniles (μm) |
花鲈脾脏外层为由结缔组织和扁平细胞组成的被膜,被膜往里为实质组织,结缔组织往内部延伸使实质组织形成脾小叶,小叶间排列紧密界限不明显。脾脏内有大量的血管和血细胞。脾实质由红髓质和白髓组成,二者分界不明显,互相穿插;红髓染色较浅,主要由红细胞组成;白髓染色较深,主要由大量的淋巴细胞和少量巨噬细胞及颗粒细胞组成;脾细胞形态多样,呈圆形、椭圆形或多角形(图 2)。盐度为0时,脾脏有较多血细胞,淋巴细胞较少(图 2.1);盐度为10时,血细胞减少,有较多的巨噬细胞,淋巴细胞聚集中心明显(图 2.2);盐度为15时,出现较多的黑色素巨噬细胞中心(图 2.3);盐度为20时,脾窦范围增大,血细胞增多(图 2.4)。盐度为30时,见部分脾细胞和淋巴细胞肿大,并出现空泡,细胞间排列较为疏松,有少许细胞溶解,差异较为明显(图 2.5)。

|
图 2 盐度对花鲈幼鱼脾脏的影响
Fig.2 The effect of salinity on the spleens of L. maculatus juveniles
1:盐度0;2:盐度10;3:盐度15;4:盐度20;5:盐度30,“↑”示淋巴细胞肿大和空泡化 BC:血细胞;E:椭圆体;EC:内皮细胞;C:脾索;G:粒细胞;L:淋巴细胞;LC:淋巴细胞中心; M:巨噬细胞;MC:黑色素巨噬细胞中心;S:脾窦;ST:脾小梁;SRBC:衰老红细胞 1: Salinity 0; 2: Salinity 10; 3: Salinity 15; 4: Salinity 20; 5: Salinity 30. "↑"indicate lymphocyte enlargement and vacuolation BC: Blood cell; C: Splenic cord; E: Ellipsoid; G: Granulocyte; L: Lymphocyte; LC: Lymphocytes center; M: Macrophage; MC: Melano-macrophage center; S: Splenic sinus; ST: Spleen trabecular; SRBC: Senile red blood cell |
花鲈幼鱼肌肉组织由神经、结缔组织、血管以及形状大小不规则的肌纤维组成,肌纤维由结缔组织包围成纤维束,肌纤维有许多细胞核,排列在基膜附近,椭圆形或梭形,呈蓝紫色。肌肉横切肌纤维呈多角形、椭圆形或梭形,排列较紧密,直径不均一(图 3)。盐度为0时,肌纤维排列较为疏松,多角形或长椭圆形,长径和短径较大,密度较小(表 3)。盐度为10时,肌纤维长径变小,短径变化不明显,排列较紧密,形状较规则,呈多角形,密度显著增大(图 3、表 3)。盐度为15时,肌纤维短径变小,密度增大,与S0、S10组均差异显著(P < 0.05)。盐度为20时,肌纤维椭圆形或多角形,排列疏松,纤维束间隙较大(图 3d)。盐度为30时,肌纤维长径增大,密度减小(表 3)。

|
图 3 盐度对花鲈幼鱼肌肉的影响
Fig.3 The effect of salinity on the muscle of L. maculatus juveniles
1:盐度0;2:盐度10;3:盐度15;4:盐度20;5:盐度30 BC:毛细血管;CT:结缔组织;MF:肌纤维;MN:肌细胞核;NF:神经纤维 1: Salinity 0; 2: Salinity 10; 3: Salinity 15; 4: Salinity 20; 5: Salinity 30 BC: Blood capillary; CT: Connective tissue; MF: Muscel fiber; MN: Myocyte nucleus; NF: Nerve fiber |
|
|
表 3 盐度对花鲈幼鱼肌肉测量性状的影响 Tab.3 The effect of salinity on the muscle meristic characters of L. maculatus juveniles |
该实验中各组花鲈幼鱼的存活率都较高,表明在0~30的盐度范围内,花鲈幼鱼能够保持完整的渗透调节能力,能够生长、存活。
该实验结果显示,花鲈在淡水和低盐环境中(S0和S10)其鳃丝和鳃小片较宽、形态较长,细胞饱满,排列紧密,有利于与水体充分接触,更易摄入水中的无机离子以适应低渗环境;泌氯细胞体积较小,数量很少,表现出适应低渗环境的形态特征;扁平细胞较厚,排列紧密,可能是防止离子自由渗漏(Evans et al, 2005)。随着盐度的增加,鳃丝宽度变小,鳃小片间距增大,长度变短;泌氯细胞体积显著增大。当盐度达到20时,部分鳃小片变形脱落,鳃丝上的泌氯细胞明显增多增大;盐度为30时,鳃丝宽度较大,出现断裂脱落,鳃丝上细胞排列疏松,泌氯细胞明显膨大,见溶解状态。这与鲻鱼(Mugil cephalus)、罗非鱼(Oreochromis mossambicus)(于娜等, 2012; Nolan et al, 1999)的研究结果基本一致。随着盐度的增加,花鲈幼鱼为适应环境需增加水、氧交换量以维持渗透平衡,其鳃丝的结构发生一定程度的萎缩,鳃丝细胞缩小和鳃小片间距增大,当盐度超过一定的耐受范围(盐度为30时),其鳃丝结构不同程度的脱落或分离。鳃丝上的泌氯细胞数量明显增加,胞体变大,说明其泌氯功能和细胞的代谢水平明显提高,发挥了Na+、K+、Cl-等离子代谢平衡调节作用(Prunet et al, 1994)。花鲈为维持机体渗透平衡,在不同盐度下,鳃组织发生适应性改变,过高的盐度波动会造成组织损伤。
脾脏是硬骨鱼类的免疫、造血和滤血器官,其主要免疫细胞有淋巴细胞、颗粒细胞、巨噬细胞等(王鸿鹄等, 2006;李长玲等, 2002)。各种免疫细胞具有机体免疫和吞噬的功能。低盐胁迫下,许氏平鲉(Sebastes schlegeli)脾脏组织中淋巴细胞数量增多聚集,粒细胞形态结构变化较显著(王晓杰等, 2006)。该研究结果显示,淡水环境中(盐度为0),花鲈幼鱼脾脏有较多红细胞,淋巴细胞较少,免疫能力较弱。随着盐度增加,在盐度为10和15的环境中,淋巴细胞增加,出现明显的淋巴细胞聚集中心,此外,巨噬细胞增多,黑色素巨噬细胞聚集,颗粒细胞增大,推测是由于盐度胁迫使其产生适应性免疫反应,增强脾脏免疫功能。在盐度为30时,脾细胞和部分淋巴细胞出现肿大、空泡化现象,可能是由于高盐度长时间胁迫使花鲈幼鱼免疫细胞受损,导致整体细胞功能减弱;严重的部分细胞溶解,黑色素巨噬细胞聚集中心减少,推测高盐度胁迫使巨噬细胞的结构受损,阻碍黑色素巨噬细胞的集结发育,黑色素巨噬细胞中心数量减少。高盐度的水环境和较大的盐度变化会对花鲈幼鱼的生长发育产生不利影响。
肌肉的组织结构主要由构成肌肉的肌纤维数量、肌纤维的直径大小、长度以及肌原纤维决定,而肌纤维直径是描述肌肉特征的重要参数。研究表明,营养水平、运动方式、环境因子等直接影响肌纤维的发育(郭琼林等, 1993; Johnston et al, 2000、2003)。研究结果显示,随着盐度的增加,花鲈幼鱼肌纤维长径和短径均有先减小后增加的趋势,单位面积肌纤维数量则呈先增加后减小的趋势,差异显著(P < 0.05)。研究表明,肌纤维数目在出生前已固定,因此,在出生后的生长发育中主要是肌纤维直径与长度的增加(Stickland et al, 1975)。肌纤维密度反映了肌肉增生速率的大小,推测是鱼类增长速度在不同盐度下差异的原因之一(关文静等, 2008)。本研究在不同盐度条件下,花鲈幼鱼的肌肉组织结构产生差异,随着肌纤维增粗,肌肉间结缔组织与脂肪组织的增加,肌纤维密度下降,该结果可为解析其生长调控机理提供参考。
Chen SX, Wang PF, Ou YJ, et al. Acute and chronic hypoxia effect on gills of golden pompano (Trachinotus ovatus). South China Fisheries Science, 2017, 13(1): 124-130 [陈世喜, 王鹏飞, 区又君, 等. 急性和慢性低氧胁迫对卵形鲳鲹鳃器官的影响. 南方水产科学, 2017, 13(1): 124-130 DOI:10.3969/j.issn.2095-0780.2017.01.016] |
Ern R, Esbaugh AJ. Effects of salinity and hypoxia-induced hyperventilation on oxygen consumption and cost of osmoregulation in the estuarine red drum (Sciaenops ocellatus). Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology, 2018, 222: 52-59 DOI:10.1016/j.cbpa.2018.04.013 |
Evans DH, Piermarini PM, Choe KP. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiological Reviews, 2005, 85(1): 97-177 |
Gibbons TC, McBryan TL, Schulte PM. Interactive effects of salinity and temperature acclimation on gill morphology and gene expression in threespine stickleback. Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology, 2018, 221: 55-62 DOI:10.1016/j.cbpa.2018.03.013 |
Guan WJ, Zhu YF, Chen ZD. Muscle quality in fish related to characteristics of muscular fibers. Fisheries Science, 2008, 27(2): 101-104 [关文静, 朱艺峰, 陈芝丹. 鱼类肌纤维特性与鱼肉品质关系. 水产科学, 2008, 27(2): 101-104 DOI:10.3969/j.issn.1003-1111.2008.02.014] |
Guo QL, Lu QZ. Observation on the development of blood cells in kidney and spleen of grass carp. Acta Hydrobiologica Sinica, 1993, 17(1): 40-45 [郭琼林, 卢全章. 草鱼肾脏和脾脏血细胞发育过程的观察. 水生生物学报, 1993, 17(1): 40-45] |
He YF, Wang LM, Liu N, et al. Effects of acute salinity stress and chronic salinity acclimatization on survival and muscle component of Malaysia red tilapia. Journal of Dalian Ocean University, 2016, 31(3): 280-284 [何燕富, 王兰梅, 刘念, 等. 急性盐度胁迫和慢性盐度驯化对马来西亚红罗非鱼存活及肌肉组分的影响. 大连海洋大学学报, 2016, 31(3): 280-284] |
Hu XW, Shangguan JB, Li ZB, et al. Effects of dietary chitosan oligosaccharide on the performance, digestion and serum biochemical indexs of the juvenile Japanese seabass (Lateolabrax japonicus). Acta Oceanologica Sinica, 2018, 40(2): 69-76 [胡晓伟, 上官静波, 黎中宝, 等. 饲料中添加壳寡糖对花鲈(Lateolabrax japonicus)幼鱼的生长、消化和血清生化指标的影响. 海洋学报, 2018, 40(2): 69-76] |
Johnston IA, Alderson R, Sandham C, et al. Muscle fibre density in relation to the colour and texture of smoked Atlantic salmon (Salmo salar L.). Aquaculture, 2000, 189(3-4): 335-349 DOI:10.1016/S0044-8486(00)00373-2 |
Johnston IA, Manthri S, Alderson R, et al. Freshwater environment affects growth rate and muscle fibre recruitment in seawater stages of Atlantic salmon (Salmo salar L.). Journal of Experimental Biology, 2003, 206(8): 1337-1351 DOI:10.1242/jeb.00262 |
Li CL, Cao FJ, Liu CW, et al. The microstructure of spleen and head kidney in Plectorhynchus cinctus. Marine Science Bulletin, 2002, 21(2): 30-35 [李长玲, 曹伏君, 刘楚吾, 等. 花尾胡椒鲷脾脏和头肾显微结构的观察. 海洋通报, 2002, 21(2): 30-35 DOI:10.3969/j.issn.1001-6392.2002.02.005] |
Li FX, Wang PF, Yan LL, et al. Molecular cloning and expression pattern analysis of irak4 in sea perch (Lateolabrax maculatus). South China Fisheries Science, 2018, 14(5): 70-79 [李富祥, 王鹏飞, 闫路路, 等. 花鲈irak4基因cDNA的克隆与表达分析. 南方水产科学, 2018, 14(5): 70-79] |
Luo HZ, Li WY, Fu RB, et al. The effects of salinity on the growth of juvenile Eleutheronema tetradactylum and Na+/K+- ATP enzyme. Progress in Fishery Sciences, 2015, 36(2): 94-99 [罗海忠, 李伟业, 傅荣兵, 等. 盐度对四指马鲅(Eleutheronema tetradactylum)幼鱼生长及其鳃丝Na+/K+- ATP酶的影响. 渔业科学进展, 2015, 36(2): 94-99] |
Nolan DT, Op't Veld RLJM, Balm PHM, et al. Ambient salinity modulates the response of the tilapia, Oreochromis mossambicus (Peters), to net confinement. Aquaculture, 1999, 177(1-4): 297-309 DOI:10.1016/S0044-8486(99)00093-9 |
Pan YY, Gu XY, Zhang DM, et al. Effects of fasting and refeeding on Lateolabrax japonicus intestinal microflora diversity. Journal of Fisheries of China, 2016, 40(5): 776-784 [潘艳艳, 顾晓英, 张德民, 等. 饥饿及恢复喂食对花鲈肠道菌群多样性的影响. 水产学报, 2016, 40(5): 776-784] |
Prunet P, Pisam M, Claireaux JP, et al. Effects of growth hormone on gill chloride cells in juvenile Atlantic salmon (Salmo salar). American Journal of Physiology, 1994, 266(2): 850-857 |
Qin GX, Wei Q, Yu JQ. Histological characterization muscular and gill of Gymnocypris przewalskii. Journal of Qinghai University (Natural Science), 2010, 28(2): 4-7 [秦桂香, 魏青, 余家庆. 青海湖裸鲤肌肉和鳃组织结构特征. 青海大学学报(自然科学版), 2010, 28(2): 4-7 DOI:10.3969/j.issn.1006-8996.2010.02.002] |
Shivakumar K, Jayaraman J. Salinity adaptation in fish: Interaction of thyroxine with fish gill mitochondria. Archives of Biochemistry and Biophysics, 1986, 245(2): 356-362 DOI:10.1016/0003-9861(86)90226-2 |
Shui C, Shi Y, Hua X, et al. Serum osmolality and ions, and gill Na+/K+-ATPase of spottedtail goby Synechogobius ommaturus (R.) in response to acute salinity changes. Aquaculture and Fisheries, 2018, 3(2): 79-83 DOI:10.1016/j.aaf.2018.03.002 |
Stickland NC, Widdowson EM, Goldspink G. Effects of severe energy and protein deficiencies on the fibres and nuclei in skeletal muscle of pigs. British Journal of Nutrition, 1975, 34(3): 421-428 DOI:10.1017/S0007114575000487 |
Wang HH, Li CL, Cao FJ, et al. Microstructure of head-kindey and spleen in Lutjanus sanguineus. Journal of Oceanography in Taiwan Strait, 2006, 25(3): 360-367 [王鸿鹄, 李长玲, 曹伏君, 等. 红笛鲷头肾和脾脏显微结构的观察. 台湾海峡, 2006, 25(3): 360-367 DOI:10.3969/j.issn.1000-8160.2006.03.009] |
Wang XJ, Zhang XM, Jiang M. Salinity stress on the ultrastructure of gill, head kidney and spleen of rockfish (Sebastes schlegeli). Periodical of Ocean University of China, 2006, 36(S1): 85-90 [王晓杰, 张秀梅, 姜明. 盐度胁迫对许氏平鲉鳃、头肾、脾脏超微结构的影响. 中国海洋大学学报, 2006, 36(S1): 85-90] |
Yu N, Li JE, Ou YJ, et al. Structural changes in gill and kidney of juvenile grey mullet under different salinity. Ecological Science, 2012, 31(4): 424-428 [于娜, 李加儿, 区又君, 等. 不同盐度下鲻鱼幼鱼鳃和肾组织结构变化. 生态科学, 2012, 31(4): 424-428 DOI:10.3969/j.issn.1008-8873.2012.04.014] |
Zhang TT, Chen XR, Liang MQ, et al. Effects of different protein hydrolysates on growth performance and non- specific immunity of Japanese seabass (Lateolabrax japonicus). Progress in Fishery Sciences, 2017, 38(3): 96-105 [张婷婷, 陈效儒, 梁萌青, 等. 不同蛋白水解物对花鲈(Lateolabrax japonicus)生长性能及非特异性免疫的影响. 渔业科学进展, 2017, 38(3): 96-105] |
Zhang XY, Wen HS, Zhang KQ, et al. Analysis of the isotonic point and effects of seawater desalination on the Na+/K+/Cl- concentration, Na+-K+-ATPase activity and relative gene expressions in Lateolabrax japonicus. Journal of Fisheries of China, 2018, 42(8): 1199-1208 [张晓燕, 温海深, 张凯强, 等. 花鲈等渗点分析及海水淡化对Na+/K+/Cl-浓度、Na+-K+-ATP酶活性及基因表达的影响. 水产学报, 2018, 42(8): 1199-1208] |