



2. 潍坊工程职业学院 山东 潍坊 262500;
3. 青岛海洋科学与技术试点国家实验室海洋生态与环境科学功能实验室 山东 青岛 266071
2. Weifang Engineering Vocational College, Weifang, Shandong 262500, China;
3. Laboratory for Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, Shandong 266071, China
江豚隶属于鲸目(Cetacea)、齿鲸亚目(Odontoceti)、鼠海豚科(Phocoenidae)、江豚属(Neophocaena),包括印度洋–太平洋江豚(N. phocaenoides,下文简称印太江豚)和窄脊江豚(N. asiaeorientalis) 2个种,其中,窄脊江豚又分为2个亚种,分别为长江江豚(N. a. asiaeorientalis)和东亚江豚(N. a. sunmeri)(Jefferson et al, 2011)。江豚主要分布在西太平洋和印度洋沿海水域,在中国见于渤海、黄海、东海、南海和长江等水域,在长江甚至能上溯到宜昌一带。随着社会经济的发展,水生野生动物的生存空间被严重挤压,常见的鲸、海豚等动物数量逐渐减少,甚至消失。江豚全部物种已被列入《世界自然保护联盟》(IUCN)国际野生动物红皮书和《濒危野生动植物种国际贸易公约》,在2021年2月5日新调整的《国家重点保护野生动物名录》中,长江江豚被列为国家一级保护野生动物东亚亚种和印太江豚为国家二级保护动物。长江江豚是全世界唯一已知的江豚淡水种群,同时也是国内研究最多的亚种种群。张先锋等(1993)、于道平等(2001)和肖文等(2000)等通过历史数据、生态考察及数学模型等方法估算长江江豚种群的数量。张先锋等(1993)运用漩涡模型(Vortex 7.3)分析了长江江豚的种群生存力。杨光等(1998)编制了江豚长江种群的静态生命表,计算其种群的内禀增长能力,认为在未来时间内,江豚长江种群会出现负增长。学者们在长江江豚行为学、繁殖生物学、遗传学、生物声学以及生境选择、保护生物学等方面都开展了一定的研究(华元渝等, 1994; 束家宽等, 2008; 郝玉江等, 2006; 杨光等, 1997; 王丁, 1996; 于道平, 2003; 于道平等, 2005)。此外,国家对长江江豚的种群数量组成进行了多次专项调查和评估,如2012年长江淡水豚考察报告的长江江豚种群数量为1040头(Mei et al, 2014);2017年考察报告的长江江豚种群数量为1020头(Huang et al, 2020)。
与长江江豚相比,海洋江豚的研究较少。王丕烈(1979、1984a)、王丕烈等(1989)和董金海等(1993)研究了江豚在黄渤海的分布和迁移情况,比较了北方江豚和南方江豚的差异。王丕烈(1992)报道了黄渤海江豚的外形和骨骼特征,还对江豚的生殖期进行了初步分析。高安利等(1995a、1995b、1995c)分别对中国水域江豚的颅骨、颅后骨和外形进行了比较。黄宗国等(2000)研究了福建沿海的江豚,发现该海域既有东亚江豚,也有印太江豚。鹿志创等(2016)应用碳氮稳定同位素技术研究江豚食性,认为江豚的食物以鱼类为主,喜食顺序为中上层鱼类 > 中下层鱼类 > 底层鱼类,也捕食头足类、虾类和蟹类等。
党的“十八大”将生态文明建设纳入五位一体的国家战略,生态大保护和高质量发展成为发展的主要态势,也为水生生物保护提供了政策依据。农业农村部“渔业第十三个五年计划”等规划都将海洋珍稀、濒危动物保护和研究列为重点工作。在此背景下,珍稀濒危水生野生动物,尤其是江豚的研究备受关注,新闻媒体的报道也日益增多,为此有必要对以往的工作加以总结,为未来的工作提供借鉴。鉴于长江江豚的研究众多,且不乏研究进展一类的论文,本文重点对海洋江豚的研究加以归纳、总结,以期为海洋江豚研究提供帮助。
1 形态与分类江豚的命名最初由Cuvier(1829)将采自印度的标本命名为Delphinus phocaenoides。后来根据采样地域的差异,出现了诸多命名,如Delphinus melas (Temminck et al, 1844),Delphinapterus molagon (Owen, 1866)和Neomeris kurrachiensis (Murray, 1884)等。Gray(1846)为江豚建立新属Neomeris。由于存在同种异名、同名异属的问题,根据命名法规,江豚学名后被改为Neophocaena phocaenoides Cuvier 1829。
Pilleri等(1972)根据长江的江豚标本同印度、巴基斯坦的江豚形态差异提出江豚属有2个种,将分布于中国沿海和长江的江豚另命名为新种Neomeris asiaeorientalis。后来,Pilleri等(1975)又根据印度、巴基斯坦产江豚标本、长江产江豚标本和日本产江豚标本三者之间的形态差异,提出江豚属包括3个种,即分布于印度洋巴基斯坦、印度沿岸的江豚为Neophocacna phocaenoides,分布于中国沿岸和长江中的江豚为Neophocaena asiaeorientalis,分布于日本沿岸的江豚为新种Neophocaena sunameri。周开亚等(1980)也支持了Pilleri等(1975)的观点。
王丕烈(1984a、1984b)对黄渤海江豚、长江口崇明岛水域江豚及南方海域的江豚进行了调查,通过对比研究首次发现,中国南北各海区的江豚形态学上存在差异,以此提出中国南北海区和长江流域分布的江豚属于不同的地理种群。之后,又对渤海、黄海、东海、南海、长江和钱塘江的江豚进行了外形和骨骼对比研究,提出不同水域江豚外形和骨骼存在的差异,指出分布于南海的江豚与印度、巴基斯坦的江豚外形、骨骼无大差异,分布于黄渤海的江豚与日本沿海的江豚外形、骨骼无大差异,认为中国产江豚应包括3个亚种,即主要分布于南海的指名亚种(N. p. phocaenoides),主要分布于东海、长江、钱塘江的扬子亚种(N. p. asiaeorientalis)和主要分布于黄渤海的北方亚种(N. p. sunameri)。
高安利等(1995a、1995b、1995c)选用了35项外形测量指标,对1974—1993年间在中国长江和大陆沿岸海域采集各年岭段的江豚标本226号进行测量和分析发现,黄海种群的吻长相对最长,南海种群的吻长相对最短。长江种群的体围比较大,黄海和南海种群的体围较小。南海种群的眼裂高、眼裂长和鳍肢宽最大。在种群之间差异最大、可作为分类鉴别特征的指标是疣粒区宽、疣粒列数和背脊高。根据疣粒区宽度和疣粒列数将南海种群与其他2个种群分开;通过背脊高度和疣粒列数将长江种群与黄海种群分开,认为江豚3个亚种中江豚指名亚种的背脊始于体背后部,体背部疣粒区宽为48~120 mm,疣粒列数为10~ 14列,分布稀疏;江豚北方亚种的背脊始自体长之半或其前,高通常在16 mm以上,疣粒区宽为3~12 mm,疣粒列数为1~9列,排列紧密;江豚长江亚种的背脊始自体长之半或其前,高通常不超过15 mm,疣粒区宽为2~8 mm,疣粒列为数2~5列。
由于形态上存在较明显的差别,但因缺乏足够的标本对比或系统的研究,长期以来对江豚的分类争议不断。直到2011年,Jefferson等(2011)根据对之前十几年的最新的形态学和分子生物学资料进行了详细研究,发现所有的江豚标本仅存在宽背部疣粒区和非常窄背部疣粒区2种情况,不存在重叠的情况,而且它们之间在1.8万年前已经出现生殖隔离。因此认为,有足够的证据证实江豚存在2个独立物种,即印太江豚(Indo-Pacific finless porpoise, Neophocaena phocaenoides)和窄脊江豚(narrow-ridged finless porpoise, N. asiaeorientalis),其中,窄脊江豚包含2个亚种,分别是长江江豚(Yangtze finless porpoise, N. a. asiaeorientalis)和东亚江豚(East Asian finless porpoise, N. a. sunameri) (Jefferson et al, 2011)。
Zhou等(2018)发表了中美科研团队关于长江江豚分子生物学的最新研究成果。研究发现,一些与水盐代谢和渗透调节相关的基因在长江江豚中出现了显著的正选择,提示它们为适应与海洋高盐环境完全不同的淡水低渗环境,已经出现了适应性进化,可更好地在淡水环境下维持体内的水盐平衡。而这种显著的遗传分化与适应性进化提示,它们之间缺乏基因交流而出现了生殖隔离。因此,长江江豚被认定为独立物种。
2 种群与分布关于江豚的记载最早见于东汉时期,《说文解字》记载,“ 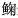
时至今日,人们对江豚的分布已非常清楚。印太江豚主要分布于印度洋和东南亚的浅水区,即西起波斯湾,东至台湾海峡;窄脊江豚主要分布于太平洋西海岸的温带水域,即南起我国台湾海峡,北至韩国和日本中部,以及我国长江中下游水域和鄱阳湖、洞庭湖(Jefferson et al, 2011)。在我国沿海、长江和日本、韩国、越南、马来西亚、巴基斯坦、印度等国家的沿岸海域均有江豚分布。早期研究把江豚作为1个物种记载,例如,陈万青(1978)的描述:江豚分布很广,自非洲好望角,至印度洋、印度、巴基斯坦,整个东南亚,向北到日本、朝鲜沿岸都有。我国沿海分布亦很普遍。它们通常栖于咸、淡水交界处或潮流冲击的地方,所以大、小河川的河口及下游一带常有发现,如长江、湘江、洞庭湖都有,甚至可沿长江上溯1000多km达湖北宜昌一带。王丕烈(1984a、1984b)报道,江豚分布于我国南海、东海、黄河和渤海,可达41ºN、122ºE。江豚在日本的分布也非常广泛,在日本海、濑户内海和太平洋沿岸都有分布(Kasuya et al, 1979)。
随着沿海国家、地区的研究不断深入,江豚的分布规律也逐步清楚。Shirakihara等(1994)调查发现,九州西部沿岸水域江豚偏向分布于水深小于50 m、离岸2~3 n mile的水域。Zhang等(2004)调查发现,韩国西海岸沿岸目击的江豚有超过80%分布在离岸15 n mile范围内,其中,78%的分布水深在20~50 m之间。Jefferson等(2002)调查发现,春季香港水域范围内分布约152头江豚。Smith等(2008)发现,孟加拉湾江豚(印太江豚)主要分布于较浅、盐度较低的水域。左涛等(2018)报道,江豚在渤海各海湾和中部水域都可观察到,以14:00~18:00时段、离岸15~35 km范围内较易观察到。黄河口外、秦皇岛东北侧老龙口和莱州外沿岸河口水域,江豚出现频次较多。Minton等(2011)发现,马来西亚砂劳越沿岸小型鲸类在观察到的115次中有100次是在水深小于10 m的水域,江豚的分布相比其他小型鲸类要更向外一些。董金海等(1993)研究认为,黄海江豚在春季游近沿岸,在10—11月逐渐返回越冬场,冬季只在深水域时有发现。在不同海域离岸的距离差异较大,可能主要是与水深相关,由此可见,江豚的分布与水深的关系比较密切。
由于海洋江豚的分布区域宽广,对于整个种群的数量调查非常困难,但仍不乏局部水域的评估报告,其中,日本的研究相对较多。Yoshida等(1997)采用截线调查法对有明湾(Ariake Sound)和立川湾(Tachibana Bay)进行了航空观测,估算得出,有明湾江豚种群数量为1983头[95%置信区间(CI)=1382~2847]、立川湾为1110头(95% CI=642~1920),2个水域综合评估为3093头(1.3头/km2,95% CI=2278~4201)。Amano等(2003)采取相同方法评估了日本太平洋沿岸仙台湾至东京湾江豚的数量,认为仙台湾至BOSO半岛江豚的密度和数量分别为0.502头/km2和3387头。Shirakihara等(2007)调查结果显示,日本内海131°3′~134°59′E区域内江豚数量为7572头,密度为0.506头/km2。Zhang等(2004)采取目视截线调查法对韩国西海岸水域江豚种群数量进行评估,报道2001年离岸水域江豚数量约为58, 650头(95% CI=34, 961~98, 389),2003年近岸水域约为1571头(95% CI=881~2800)。我国除长江江豚有系统的调查和种群数量评估外,海洋江豚种群数量调查仅见于南海的北部湾和渤海的报道。徐信荣等(2012)在2003—2004年完成了30个航次总航程1906 km调查,未发现江豚;2011—2012年完成71个航次总航程2976.2 km调查,发现江豚6次,遇见率为仅为0.005头/km。左涛等(2018)估算渤海江豚种群数量为7883头(变异系数CV=0.21)、3701头(CV=0.21)和3124头(CV=0.21),对应的种群密度分别为1.441、0.066和0.056头/km2。
Shirakihara等(2007)认为,日本沿海江豚有5个亚种群,1978—2000年的22年间种群总体数量下降了69%。Park等(2015)报道,2004—2005年韩国沿海江豚平均密度为0.565头/km2,近岸水域分布密度为0.638头/km2,到2011年沿海和近岸水域江豚平均密度分别降为0.122和0.151头/km2,种群数量下降近70%。
3 其他方面的研究与长江江豚的研究相比,关于海洋江豚的生物、生态、繁殖、行为、摄食、遗传多样性和声学等方面的研究少有报道。
3.1 生物学江豚基础生物学的研究是和形态学研究同步开展的。王丕烈(2012)对不同水域江豚的体长、体重进行了总结,黄渤海所获67头(雌43,雄24)江豚的平均体长为140 cm、平均体重为43.6 kg,其中,雌性最大体长为220 cm,平均体长为136.7 cm;雄性最大体长为227 cm,平均体长为150.1 cm;长江口崇明岛附近的51头(雌20,雄31)江豚的平均体长为135 cm,雌性最大体长为154 cm,平均体长为132.2 cm,雄性最大体长为170 cm,平均体长为137.6 cm;南海区的江豚指名亚种个体偏小,雄性和雌性的最大体长分别为168和165 cm。此外,通过获得江豚胎儿及1例幼体推测,黄渤海初生仔豚的体长为80~93 cm (王丕烈, 2012)。Kasuya等(1979)报道,日本沿岸江豚初生仔豚体长为77.5~83.0 cm。
3.2 生态学董金海等(1993)研究认为,海洋江豚的迁移主要与饵料鱼类的洄游分布等有关,春季随鱼类产卵群体主要分布在近岸的产卵场,而秋冬季随鱼类洄游迁移至黄海鱼类的越冬场,但这与江豚主要分布在较浅(一般在50 m等深线以浅)水域有所不同。左涛等(2018)报道,在各调查季节中,5月目击江豚的次数和头数最多,遇见率也最高,9月和11月江豚遇见率较低;5月,白天各时段都可目击到江豚,6—8月目击到江豚的时段主要在14:00—18:00,9月和11月的目击时段多在14:00以前。作者认为,江豚分布的季节差异可能与江豚食物的分布和活动有关。例如,多种鱼类被报道在5月前后洄游产卵(陈大刚, 1991; 李建生等, 2014)。需要注意的是,5月海况较好,海雾主要发生在上午,也可能对渤海江豚调查结果产生一定的影响(左涛等, 2018)。
3.3 摄食Shirakihara等(2008)通过对1987—1992年在有明湾、立川湾和野村湾(Omura Bay)收集的87头江豚胃含物的分析发现,在野村湾江豚胃含物中鱼类(虾虎鱼类和银汉鱼类)的数量最多,出现频率最高;在有明湾和立川湾江豚胃含物中头足类和鱼类(鲱科、鳀科和石首鱼科)的比重相当。鹿志创等(2016)应用碳氮稳定同位素技术对海洋江豚的食性进行了研究,分析结果显示,江豚的食物来源主要以鱼类为主,对食物种类的喜食顺序为中上层鱼类 > 中下层鱼类 > 底层鱼类 > 头足类 > 虾类 > 蟹类,其平均贡献率分别为43.9%、18.2%、13.1%、10.1%、8.8%和6.0%。江豚的营养级为4.5,高于传统胃含物分析法的研究结果。Lu等(2016)使用形态学和mtDNA条形码技术对采集自渤海、黄海和东海的共23头东亚江豚的胃含物进行了分析,结果显示,江豚的食物中包括鱼类、头足类、甲壳类、双壳贝类,其中,拉氏狼牙虾虎鱼(Odontamblyopus lacepedii)、澳洲鲐(Scomber australasicus)、小黄鱼(Larimichthys polyactis)、黄姑鱼(Nibea albiflora)和长蛇鲻(Saurida elongata)为主要食物,且存在地理间差异。Chen等(2017)使用形态学和分子生物学方法对采集自东海的122头江豚的胃含物进行了分析,结果显示,鱼类是江豚的主要食物,头足类和甲壳类是次要食物,且不同性别和不同年龄个体间存在差异。王丕烈(2012)认为,江豚的食性较广,以鱼类为主,也摄食虾类和头足类。江豚在黄渤海主要摄食青鳞鱼(Sardinella zunasi)、大银鱼(Protosalanx hyalocranius)、叫姑鱼(Johnius belengerii)、梭鱼(Liza haematocheila)、鲈(Lateolabrax japonicus)、澳洲鲐、小黄鱼、对虾(Penaeus chinensis)和头足类等;在长江口春季主要摄食大银鱼、刀鲚(Coilia nasus)等。
3.4 繁殖王丕烈(1979)对黄渤海小型齿鲸类调查发现,江豚有抚幼行为,在春季4—5月,常见仔豚用鳍肢趴伏在母豚背部,由母豚驮负游泳。由此推断,江豚的繁育期在这个时间段。王丕烈(1984b)观察发现,江豚具有护幼行为,例如,在仔豚被驱赶从母豚背后掉落后,母豚并不舍弃,会寻找到仔豚再驮负仔豚。Gao等(1993)报道,南海江豚的最低性成熟年龄雄性为4龄、雌性为5龄;黄渤海雌、雄均为5龄;南海种群出生后生长速度很快,通常在6龄时接近最大体长,8龄后体长基本不再增加,基本可全年繁殖,出生高峰在8—12月。
王丕烈(2012)根据获得的江豚胎儿、仔豚的时间段,认为黄渤海江豚的交配期从3月开始,盛期在5—7月,也有晚到8月的。从发现仔豚的时间以及获得幼体的牙齿情况推测,江豚最早2月开始产仔,盛期为3—5月。
3.5 遗传学近年来,分子生物学的方法和技术不断进步,其在海洋哺乳动物的系统进化和种群遗传研究中得到了越来越广泛的应用。杨光等(1997)运用PCR产物的银染测序技术,对中国水域的12头江豚个体的mtDNA控制区序列进行了测定,表明中国水域江豚各种群的控制区序列所定义的单倍型互不相同,无共有的单倍型;种群内的遗传分化程度远远低于种群间的遗传分化程度,提示江豚不同种群之间可能已有显著的遗传分化,种群间缺乏基因交流。此后,Yang等(2002、2003)进一步对中国水域江豚的遗传结构进行了研究,认为长江江豚、东亚江豚、印太江豚之间存在个体迁移和基因交流,其中,长江江豚的遗传多样性水平较低,与印太江豚相当,东亚江豚的核苷酸多样性和单倍型多样性最高。李东明(2005)对长江江豚和渤海江豚样品的ISSR遗传多样性进行了初步分析,发现2个种群的遗传多样性水平较低,长江江豚略高于渤海种群,二者之间的基因交流水平较高。
3.6 声学Goold等(2002)使用单水听器对香港水域江豚的回声定位信号进行记录和分析,发现该水域江豚发出2种脉冲信号,一种频带较窄,其峰值频率为142 kHz,持续时间为104 μs;另一种频带较宽,其峰值频率更高,持续时间为61 μs。Li等(2007)使用单水听器对辽东湾水域的东亚江豚的回声定位信号进行记录和分析,结果发现,其与长江江豚发出的回声定位信号参数相近:峰值频率为(121±3.78) kHz,3 dB带宽为(17.5±3.30) kHz,持续时间为(80±11.49) μs,周波数为(9±1.48)个。Akamatsu等(2008、2010)和Nakamura等(2016)采用被动声学方法对日本多处水域江豚的监测显示,声学探测主要集中在夜间,并在3月和4月达到峰值。
4 未来工作建议1988年江豚被列为国家二级保护动物。1996年长江江豚被国际自然保护联盟列为“濒危级”。目前,江豚被列入《濒危野生动植物种国际贸易公约》(CITES Appendix Listing)附录Ⅰ物种;被列入《中国物种红色名录》濒危级(EN);2017年,IUCN评估等级为“濒危(EN)”。2017年全国两会期间,10名驻鄂全国政协委员联名提案:提升长江江豚的保护级别,将其由国家二级保护动物提升为国家一级保护动物。农业农村部长江流域渔政监督管理办公室5月在上海组织召开了“长江江豚升级为国家一级保护动物专题论证会”,与会专家经过热烈的质询与讨论,一致同意长江江豚保护等级应为一级。2021年2月5日新调整的《国家重点保护野生动物名录》,正式将长江江豚列为国家一级保护野生动物。
相比较而言,海洋江豚的保护远不及长江江豚。究其原因,一是对于海洋动物的保护意识远远落后于内陆流域,更无法与陆生动物相比。调研发现,很多渔业从业者根本不知道海洋里的江豚是国家二级保护动物,甚至有买卖现象。二是基础研究严重滞后,尽管很早已开展分类、分布的调查和研究,但时至今日仍没有系统、全面地开展海洋江豚调查,对整个江豚的种群及其数量不了解,只有局部海域的评估结果。三是海洋江豚分布范围广,我国沿海均有分布,加之我国海洋捕捞力量强,捕捞对江豚的损害、致死频见报道,但监管难度大。为更好保护海洋江豚,避免重蹈“白鱀豚”的覆辙,提出如下建议:
4.1 注重基础研究江豚的保护离不开基础研究的支持,而且基础研究不可滞后于保护,因此,建议开展如下研究:
4.1.1 海洋江豚的种群分布与数量调查已知海洋江豚有2个种,也有报道日本沿海存在江豚不同的种群,但对于海洋江豚是否存在不同种群(或地理种群)、种群多大及其分布范围多广、各种群间的关系如何,诸如此类的问题都亟待解决。
4.1.2 海洋江豚生态习性及栖息地研究海洋江豚主要分布在较浅的水域,但其机制还没有确切的科学根据。同样,在渤海,春、夏、秋季江豚目击率较高,到冬季却几乎见不到,有报道认为,冬季江豚随鱼类越冬洄游进入越冬场海域,然而,多次冬季河口江豚搁浅的事例启发我们提出问题:江豚是否都会迁移?上述问题,都需要在深入研究江豚与环境的关系,掌握江豚栖息地选择的关键影响因子后,才能有更好的答案。
4.1.3 海洋江豚的遗传学和系统进化研究生物的保护离不开对其遗传、系统进化的研究,由此也就产生了进化保护遗传学。通过对海洋江豚遗传学、系统进化的研究,有助于研究与灭绝风险相关的遗传因素,了解江豚濒危机制、物种遗传多样性等,针对性选择保护策略和方式等。但目前鲜有针对海洋江豚开展的相关研究。已有研究表明,黄海东亚江豚与南海印太江豚之间遗传结构存在显著差异(Yang et al, 2002),而印太江豚不同种群间的分化程度达到了与窄脊江豚之间的分化程度(Jia et al, 2014; Zhou et al, 2018)。小种群比大种群更易受到遗传漂变导致的遗传多样性的丢失,也更可能表现出相对较低的适合度,而濒临灭绝的风险。因而,目前急需加强对海洋江豚大尺度和小尺度间种群遗传差异的研究和比较,判断是否已出现不同的地理种群。
4.2 强化保护区建设对野生动物保护来说,设立保护区并进行严格的管理,是一种行之有效的途径。以斑海豹(Phoca largha)为例,我国海域斑海豹的历史最高纪录为8000头左右,由于种群资源遭到破坏,斑海豹数量一直处于较低水平,最低时约为1200头(《斑海豹保护行动计划(2017—2026年)》)。1983年,斑海豹被辽宁省政府列为省级保护动物;1988年,斑海豹被列为国家二级重点保护动物;1992年,大连市政府批准成立了市级斑海豹自然保护区,1997年12月,经国务院批准晋升为国家级斑海豹自然保护区。经过十几年的保护区建设和保护,2006年和2007年斑海豹调查结果约为2000头(《斑海豹保护行动计划(2017—2026年)》),取得了明显成效。因此,海洋江豚保护区建设需尽早提上日程,以基础调查、研究结果为依据进行规划和建设。建议农业农村部牵头制定《海洋江豚研究保护十年计划》,国家林业局牵头启动海洋江豚保护区规划和建设工作。
4.3 注重保护技术研发健全的法律法规、保护区建设以及保护组织等,都可以禁止或限制对野生动物的直接伤害。研究证实,船舶噪声对鲸类的听觉有影响,进而影响其交流、捕食和逃避等;环境污染会造成鲸类生理变化;捕捞误捕造成死亡等。因此,有必要针对性开展如逃逸装置、规避设施等技术研发。
4.4 加强宣传教育我国颁布的《中华人民共和国野生动物保护法》第八条规定:各级人民政府应当加强野生动物保护的宣传教育和科学知识普及工作,鼓励和支持基层群众性自治组织、社会组织、企业事业单位、志愿者开展野生动物保护法律法规和保护知识的宣传活动。教育行政部门和学校应当对学生进行野生动物保护知识教育。新闻媒体应当开展野生动物保护法律法规和保护知识的宣传,对违法行为进行舆论监督。野生动物保护宣传教育,是提高民众保护意识的重要手段。
AKAMATSU T, NAKAMURA K, KAWABE R, et al. Seasonal and diurnal presence of finless porpoises at a corridor to the ocean from their habitat. Marine Biology, 2010, 157(8): 1879-1887 DOI:10.1007/s00227-010-1459-8 |
AKAMATSU T, NAKAZAWA I, TSUCHIYAMA T, et al. Evidence of nighttime movement of finless porpoises through Kanmon Strait monitored using a stationary acoustic recording device. Fisheries Science, 2008, 74(5): 970-975 DOI:10.1111/j.1444-2906.2008.01614.x |
AMANO M, NAKAHARA F, HAYANO A, et al. Abundance estimate of finless porpoises off the Pacific coast of eastern Japan based on aerial surveys. Mammal Study, 2003, 28(2): 103-110 DOI:10.3106/mammalstudy.28.103 |
CHEN DG. Fishery ecology in the Bohai and Yellow Seas. Beijing: China Ocean Press, 1991 [陈大刚. 黄渤海渔业生态学. 北京: 海洋出版社, 1991]
|
CHEN W. Handbook of marine mammals. Beijing: Science Press, 1978 [陈万青. 海兽检索手册. 北京: 科学出版社, 1978]
|
CHEN B, WANG L, WANG H, et al. Finless porpoises (Neophocaena asiaeorientalis) in the East China Sea: Insights into feeding habits using morphological, molecular, and stable isotopic techniques. Canadian Journal of Fisheries and Aquatic Sciences, 2017, 74(10): 1628-1645 DOI:10.1139/cjfas-2016-0119 |
CUVIER G. Le regne animal distribue d'apres son organsation, pour servir de base a l'histoire naturelle des animaux et d'introduction a l'anatomie comparee. Paris: Chez Deterville Libraire, 1829
|
DONG J H, WANG G J, XIAO Z Z. Migration and population difference of the finless porpoise in China. Marine Sciences, 1993(5): 42-45 [董金海, 王广洁, 肖志忠. 黄、渤海及东海江豚的洄游和种群差异. 海洋科学, 1993(5): 42-45] |
GAO A L, ZHOU K Y. Geographical variation of external measurements and three subspecies of Neophocaene Phocaenoides in Chinese waters. Acta Theriologica Sinica, 1995a, 15(2): 81-92 [高安利, 周开亚. 中国水域江豚外形的地理变异和江豚的三亚种. 兽类学报, 1995a, 15(2): 81-92] |
GAO A L, ZHOU K Y. Geographical variations of postcranial skeleton among the populations of Neophocaene in Chinese waters. Acta Theriologica Sinica, 1995b, 15(4): 246-253 [高安利, 周开亚. 中国水域江豚颅后骨骼的地理变异. 兽类学报, 1995b, 15(4): 246-253] |
GAO A L, ZHOU K Y. Geographical variations of skull among the populations of Neophocaene in Chinese waters. Acta Theriologica Sinica, 1995c, 15(3): 161-169 [高安利, 周开亚. 中国水域江豚颅骨的地理变异. 兽类学报, 1995c, 15(3): 161-169] |
GAO A L, ZHOU K Y. Growth and reproduction of the three populations of finless porpoise, Neophocaena phocaenoides, in Chinese waters. Aquatic Mammals, 1993, 19(1): 3-12 |
GOOLD J C, JEFFERSON T A. Acoustic signals from free-ranging finless porpoises (Neophocaena phocaenoides) in the waters around Hong Kong. Raffles Bulletin of Zoology, 2002, 10: 131-139 |
GRAY J E. On the cetaceous animals. In: RICHARDSON J, GRAY J E, eds. The zoology of the voyage of H.M.S. Erebus and Terror, under the command of Captain Sir James Clark Ross, R. N., F. R. S., during the years 1839 to 1843. London: Longman, 1846
|
HAO Y J, WANG D, ZHANG X F. Review on breeding biology of Yangtze finless porpoise (Neophocaene Phocaenoides asiaeorientalis). Acta Theriologica Sinica, 2006, 26(2): 191-200 [郝玉江, 王丁, 张先锋. 长江江豚繁殖生物学研究概述. 兽类学报, 2006, 26(2): 191-200 DOI:10.3969/j.issn.1000-1050.2006.02.013] |
HUA Y Y, XIANG C S, DONG M, et al. Study on the sexual and feeding behavior of the captured black finless porpoise (Neophocaene Phocaenoides) in Yangtze River. Resources and Environment in the Yangtze Valley, 1994, 3(2): 141-146 [华元渝, 项澄生, 董明, 等. 长江江豚的交配行为和摄食行为的研究. 长江流域资源与环境, 1994, 3(2): 141-146] |
HUANG Z G, LIU W H, ZHENG C X, et al. Finless porpoises (Neophocaene phocaenoides) in the southern coastal waters of Fujian, China. Acta Oceanologica Sinica, 2000, 22(5): 100-105 [黄宗国, 刘文华, 郑成兴, 等. 福建南部沿海的江豚. 海洋学报, 2000, 22(5): 100-105 DOI:10.3321/j.issn:0253-4193.2000.05.013] |
HUANG J, MEI Z G, CHEN M, et al. Population survey showing hope for population recovery of the critically endangered Yangtze finless porpoise. Biological Conservation, 2020, 241: 108315 DOI:10.1016/j.biocon.2019.108315 |
JEFFERSON T A, HUNG S K, LAW L, et al. Distribution and abundance of finless porpoises in the Hong Kong and adjacent waters of China. Raffles Bulletin of Zoology, 2002, 10: 43-55 |
JEFFERSON T A, WANG J Y. Revision of the taxonomy of finless porpoises (genus Neophocaena): The existence of two species. Journal of Marine Animals and Their Ecology, 2011, 4(1): 3-16 |
JIA K T, LIN W Z, GUI D, et al. Molecular evidence reveals the distinctiveness of Indo-Pacific finless porpoises (Neophocaena phocaenoides) in the Pearl River Estuary and insights into genus Neophocaena's origin. Marine Biology, 2014, 161(8): 1919-1930 DOI:10.1007/s00227-014-2474-y |
KASUYA T, KUREHA K. The population of finless porpoise in the inland Sea of Japan. Scientific Reports of the Whales Research Institute, 1979, 31: 1-46 |
LI D M. Analysis on genetic diversity and serum vitamin concentration of different finless porpoise (Neophocaena phocaenoides) populations. Masterxs Thesis of Nanchang University, 2005 [李东明. 不同江豚群体遗传多样性及其血清维生素含量. 南昌大学硕士研究生学位论文, 2005]
|
LI J S, HU F, YAN L P, et al. The fecundity of chub mackerel (Scomber japonicus) spawning stocks in the central East China Sea. Progress in Fishery Sciences, 2014, 35(6): 10-15 [李建生, 胡芬, 严利平, 等. 东海中部日本鲭(Scomber japonicus)产卵群体繁殖力特征. 渔业科学进展, 2014, 35(6): 10-15] |
LI S, WANG D, WANG K, et al. Echolocation click sounds from wild inshore finless porpoise (Neophocaena phocaenoides sunameri) with comparisons to the sonar of riverine Np. asiaeorientalis.. Journal of the Acoustical Society of America, 2007, 121(6): 3938-3946 DOI:10.1121/1.2721658 |
LU Z C, TIAN J S, WANG Z H, et al. Using stable isotope technique to study feeding habits of the finless porpoise (Neophocaene asiaeorientalis spp. sunameri). Acta Ecologica Sinica, 2016, 36(1): 69-76 [鹿志创, 田甲申, 王召会, 等. 应用碳氮稳定同位素技术研究江豚(Neophocaene asiaeorientalis spp. sunameri)食性. 生态学报, 2016, 36(1): 69-76] |
LU Z C, XU S Y, SONG N, et al. Analysis of the diet of finless porpoise (Neophocaena asiaeorientalis sunameri) based on prey morphological characters and DNA barcoding. Conservation Genetics Resources, 2016, 8(4): 523-531 DOI:10.1007/s12686-016-0575-2 |
MEI Z G, ZHANG X Q, HUANG S L, et al. The Yangtze finless porpoise: On an accelerating path to extinction?. Biological Conservation, 2014, 172: 117-123 DOI:10.1016/j.biocon.2014.02.033 |
MINTON G, PETER C, TUEN A A. Distribution of small cetaceans in the nearshore waters of Sarawak, east Malaysia. Raffles Bulletin of Zoology, 2011, 59(1): 91-100 |
MURRAY J A. A contribution to the knowledge of the marine fauna of Kurrachee. Annals and Magazine of Natural History, 1884, 13(77): 348-352 DOI:10.1080/00222938409459252 |
NAKAMURA K, HIRAKAWA Y, KAMEZAKI N, et al. Acoustic presence of finless porpoises around the Kansai International Airport. Techno-Ocean, 2016, 342-345
|
OWEN F R S. On some Indian Cetacea collected by Walter Elliot, Esq. Transactions of the Zoological Society of London, 1866, 6: 17-47 DOI:10.1111/j.1096-3642.1866.tb00570.x |
PARK K J, SOHN H, AN Y R, et al. A new abundance estimate for the finless porpoise Neophocaena asiaeorientalis on the West Coast of Korea: An indication of population decline. Fisheries and Aquatic Sciences, 2015, 18(4): 411-416 DOI:10.5657/FAS.2015.0411 |
PILLERI G, GIHR M. Contribution to the knowledge of cetaceans of Pakistan with particular reference to the genera Neomeris, Sousa, Delphinus and Tursiops and description of a new Chinese porpoise (Neomeris asiaeorientalis). Investigations on Cetacea, 1972, 4: 107-162 |
PILLERI G, GIHR M. On the taxonomy and ecology of the finless black porpose, Neophocaena (Cetacea, Delphinidae). Mammalia, 1975, 39(4): 657-673 |
SHIRAKIHARA K, SHIRAKIHARA M, YAMAMOTO Y. Distribution and abundance of finless porpoise in the Inland Sea of Japan. Marine Biology, 2007, 150(5): 1025-1032 DOI:10.1007/s00227-006-0363-8 |
SHIRAKIHARA M, SEKI K, TAKEMURA A, et al. Food habits of finless porpoises Neophocaena phocaenoides in Western Kyushu, Japan. Journal of Mammalogy, 2008, 89(5): 1248-1256 DOI:10.1644/07-MAMM-A-264.1 |
SHIRAKIHARA M, SHIRAKIHARA K, TAKEMURA A. Distribution and seasonal density of the finless porpoise Neophocaena phocaenoides in the coastal waters of western Kyushu, Japan. Fisheries Science, 1994, 60(1): 41-46 DOI:10.2331/fishsci.60.41 |
SHU J K, JIANG W H, ZHENG B Y, et al. Diurnal behavior and time budget of a Yangtze finless porpoise (Neophocaene phocaenoides asiaeorientalis) mother during lactation. Acta Theriologica Sinica, 2008, 28(1): 20-27 [束家宽, 蒋文华, 郑邦友, 等. 长江江豚哺乳期母豚昼间行为和时间的分配. 兽类学报, 2008, 28(1): 20-27 DOI:10.3969/j.issn.1000-1050.2008.01.004] |
SMITH B D, AHMED B, MOWGLI R M, et al. Species occurrence and distributional ecology of nearshore cetaceans in the Bay of Bengal, Bangladesh, with abundance estimates for Irrawaddy dolphins Orcaella brevirostris and finless porpoises Neophocaena phocaenoides. Journal of Cetacean Research and Management, 2008, 10(1): 45-58 |
TEMMINCK C J, SCHLEGEL H. Les mammiferes marins. In: TEMMINCK C J, ed. Apercu general et specifique sur les mammiferes qui habitant le Japon et les Iles qui en dependent. Leiden: A. Arnz & Comp, 1844
|
WANG D. A preliminary study on sound and acoustic bebavior of the Yangtze River finless porpoise, Neophocaene Phocaenoides. Acta Hydrobiologica Sinica, 1996, 20(2): 127-133 [王丁. 长江江豚声信号及其声行为的初步研究. 水生生物学报, 1996, 20(2): 127-133] |
WANG P L, XIANG F C, LIU X S. A study on finless porpoise from Bohai Sea and Yellow Sea. Fisheries Science, 1989, 8: 17-22 [王丕烈, 项福椿, 刘锡山. 黄渤海江豚的调查研究. 水产科学, 1989, 8(4): 17-22] |
WANG P L. Chinese cetaceans. Beijing: Chemical Industry Press, 2012 [王丕烈. 中国鲸类. 北京: 化学工业出版社, 2012]
|
WANG P L. Distribution of cetaceans off the coast of China. Chinese Journal of Zoology, 1984a(6): 52-56 [王丕烈. 中国近海鲸类的分布. 动物学杂志, 1984a(6): 52-56] |
WANG P L. Distribution, ecology and resource conservation of finless porpoise along Chinese waters. Transactions of Liaoning Zoological Society, 1984b, 5(1): 105-110 [王丕烈. 中国近海江豚分布、生态和资源保护. 辽宁动物学会会刊, 1984b, 5(1): 105-110] |
WANG P L. Study on the cetaceans of Bohai and Yellow Seas. Chinese Journal of Zoology, 1979(2): 31-33, 65 [王丕烈. 黄渤海产中小型齿鲸类的调查. 动物学杂志, 1979(2): 31-34, 65] |
WANG P L. The morphological characters and the problem of subspecies identifications of the finless porpoise. Fisheries Science, 1992(11): 4-8 [王丕烈. 江豚的形态特征和亚种划分问题. 水产科学, 1992(11): 4-8] |
XIAO W, ZHANG X F. A preliminary study on the population size of Yangtze finless porpoise in Poyang Lake, Jiangxi. Chinese Biodiversity, 2000, 8(1): 106-111 [肖文, 张先锋. 截线抽样法用于鄱阳湖江豚种群数量研究初报. 生物多样性, 2000, 8(1): 107-112] |
XU X R, CHEN B Y, WANG L, et al. The sympatric distribution pattern and tempo-spatial variation of Indo-Pacific humpback dolphins and finless porpoises at Shatian, Beibu Gulf. Acta Theriologica Sinica, 2012, 32(4): 325-329 [徐信荣, 陈炳耀, 王炼, 等. 北部湾沙田水域中华白海豚和江豚的同域分布格局及时空变化. 兽类学报, 2012, 32(4): 325-329] |
YANG G, LIU S, REN W K, et al. Mitochondrial control region variability of Baiji and the finless porpoises, two sympatric small cetaceans in the Yangtze River. Acta Theriologica, 2003, 48(4): 469-483 DOI:10.1007/BF03192493 |
YANG G, REN W H, ZHOU K Y, et al. Population genetic structure of finless porpoises, Neophocaena phocaenoides, in Chinese waters, inferred from mitochondrial control region sequences. Marine Mammal Science, 2002, 18(2): 336-347 DOI:10.1111/j.1748-7692.2002.tb01041.x |
YANG G, ZHOU K Y, GAO A L, et al. A study of the life table and dynamics of three finless porpoise populations in the Chinese waters. Acta Theriologica Sinica, 1998, 18(1): 1-7 [杨光, 周开亚, 高安利, 等. 江豚生命表和种群动态的研究. 兽类学报, 1998, 18(1): 1-7] |
YANG G, ZHOU K Y. Genetic variation of three populations of finless porpoise (Neophocaene phocaenoides) in Chinese waters. Acta Zoologica Sinica, 1997, 43(4): 411-419 [杨光, 周开亚. 中国水域江豚种群遗传变异的研究. 动物学报, 1997, 43(4): 411-420 DOI:10.3321/j.issn:0001-7302.1997.04.011] |
YOSHIDA H, SHIRAKIHARA K, KISHINO H, et al. A population size estimate of the finless porpoise, Neophocaena phocaenoides, from aerial sighting surveys in Ariake Sound and Tachibana Bay, Japan. Researches on Population Ecology, 1997, 39(2): 239-247 DOI:10.1007/BF02765270 |
YU D P, DONG M L, WANG J, et al. Population status of Yangtze finless porpoise in the Yangtze River section from Hukou to Nanjing. Acta Theriologica Sinica, 2001, 21(3): 174-179 [于道平, 董明利, 王江, 等. 湖口至南京段长江江豚种群现状评估. 兽类学报, 2001, 21(3): 174-179 DOI:10.3969/j.issn.1000-1050.2001.03.002] |
YU D P, WANG J, YANG G, et al. Primary analysis on habitat selection of Yangtze finless porpoise in Spring in the section between Hukou and Digang. Acta Theriologica Sinica, 2005, 25(3): 302-306 [于道平, 王江, 杨光, 等. 长江湖口至荻港段江豚春季对生境选择的初步分析. 兽类学报, 2005, 25(3): 302-306 DOI:10.3969/j.issn.1000-1050.2005.03.016] |
YU D P. Progress on conservation biology of Yangtze finless porpoise. Journal of Anhui University (Natural Science), 2003, 27(4): 98-103 [于道平. 长江江豚保护生物学研究进展. 安徽大学学报(自然科学版), 2003, 27(4): 98-103 DOI:10.3969/j.issn.1000-2162.2003.04.020] |
ZHANG C I, PARK K J, KIM Z G, et al. Distribution and abundance of finless porpoise (Neophocaena phocaenoides) in the west coast of Korea. Korean Journal of Fisheries and Aquatic Sciences, 2004, 37(2): 129-136 DOI:10.5657/kfas.2004.37.2.129 |
ZHANG X F, LIU R J, ZHAO Q Z, et al. The population of finless porpoise in the middle and lower reaches of Yangtze River. Acta Theriologica Sinica, 1993, 13(4): 260-270 [张先锋, 刘仁俊, 赵庆中, 等. 长江中下游江豚种群现状评价. 兽类学报, 1993, 13(4): 260-270] |
ZHOU K Y, PILIEN G, LI Y M. Observation of Baiji and finless porpoise from Nanjing to Taiyangzhou in the lower reaches of the Yangtze River, and physiological adaptations of Baiji porpoise to the environment. Science China, 1980(4): 363-370 [周开亚, Pilien G, 李悦民. 长江下游南京至太阳洲江段白暨豚和江豚的观察, 兼论白鱀豚对环境的某些生理适应. 中国科学, 1980(4): 363-370] |
ZHOU X M, GUANG X M, SUN D, et al. Population genomics of finless porpoises reveal an incipient cetacean species adapted to freshwater. Nature Communications, 2018, 9(1): 1276 DOI:10.1038/s41467-018-03722-x |
ZUO T, SUN J Q, SHI Y Q, et al. Primary survey of finless porpoise population in the Bohai Sea. Acta Theriologica Sinica, 2018, 38(6): 551-561 [左涛, 孙坚强, 时永强, 等. 渤海江豚种群现状初探. 兽类学报, 2018, 38(6): 551-561] |