



2. 浙江海洋大学国家海洋设施养殖工程技术研究中心 浙江 舟山 316022
2. National Engineering Research Center For Marine Aquaculture, Zhejiang Ocean University, Zhoushan 316022, China
潮间带是连接陆地和海洋的过渡区域,有丰富的生物资源,是重要的海洋湿地生态系统,主要包括泥滩、沙滩、河口等类型(林和山等, 2016; 王金宝等, 2018; 孟珊等, 2020)。由于潮间带的独特地理位置,常受到潮汐、高温、干露和缺氧等剧烈的环境变化影响,同时也容易受到人类活动的影响,因此,这类生态系统十分脆弱(刘士龙等, 2019; 金文育等, 2017),有待制定合理的保护规划。
贝类作为潮间带大型底栖生物的重要组成部分,一般栖息在潮间带沉积物中或附着于岩石上,移动能力差,对于环境的骤变较为敏感,对潮间带环境具有很好的指示作用(Blasc et al, 2008)。因此,了解潮间带贝类群落结构季节变化特征,将有助于指导潮间带生态系统保护,还能为研究潮间带贝类的生活特征提供参考。
长山列岛海域处于黄海与渤海交汇处,海岛纵列于渤海海峡中(《中国海岛志》编纂委员会, 2013),由32个岛屿组成,主岛为南长山岛和北长山岛,其岛屿底质多为岩礁或砾石。由于独特的地理位置,受到黄海和渤海两大水团的影响,黄、渤海冷水团与暖流分别在夏季和冬季有流入,渤海易受季风气候的影响(郑东等, 1983; 于非等, 2006; 石强, 2019),形成了长山列岛独特的气候特点。夏季为该群岛的旅游旺季,伴随着长山列岛养殖业和旅游业的迅猛发展,人类活动对其潮间带的干扰越来越强,对潮间带大型底栖生物群落结构造成了不同程度的影响(吴洽儿等, 2010; Niu et al, 2022)。因此,研究该岛群潮间带贝类群落组成和多样性有重要生态保护意义。
本研究对长山列岛潮间带贝类群落进行系统的四季调查,研究贝类群落分布特征和物种多样性,以期为长山列岛潮间带贝类生物多样性和环境保护提供基础资料和参考依据。
1 材料与方法 1.1 调查站点和时间2021年7月(夏季)、2021年10月(秋季)、2022年2月(冬季)和2022年5月(春季)的大潮期间,从南至北在长山列岛中选取5个较大且交通便利的岛屿进行潮间带调查。综合潮间带的面积和潮间带之间的距离,同时避开旅游景区和养殖海域等人类活动密集的场所,每个岛屿选取2个站点,共10个站点的潮间带进行贝类群落定量和定性调查。分别是南长山岛1号站点(NCD1)和2号站点(NCD2),北长山岛1号站点(BCD1)和2号站点(BCD2),砣矶岛1号站点(TJD1)和2号站点(TJD2),大钦岛1号站点(DQD1)和2号站点(DQD2),以及南隍城岛1号站点(NHD1)和2号站点(NHD2)(图 1)。在每个调查站点设置垂直于海岸的断面,每个断面分别在高、中和低3个潮区各设置垂直于断面的3个调查位点,共计30个。

|
图 1 长山列岛潮间带调查站点及位点示意图 Fig.1 Schematic diagram of the survey sites in the intertidal zone of Changshan Islands |
按照《海洋调查规范》(GB/T12763.6-2007)中的相关方法,进行样品采集、处理、鉴定和称量。样品采集时,运用手机GPS软件进行定位,每个调查位点根据潮间带垂直断面随机设置3个50 cm×50 cm的样方。由于长山列岛的底质均为岩石或砾石,则直接捡拾岩壁表面的贝类。将在每个潮区采集到的贝类样品冲洗干净后用75%酒精固定,贴上标签,注明地点和采样时间;另外,在采集地周围尽可能多采集贝类样品,同样的处理方法,带回实验室以补充样方内贝类种类上的不足。在此过程中,对样方内的贝类进行种类和数量上的粗略估数并拍照。在实验室内对样品进行分类、鉴定、计数和称量(Mettler Toledo, PL4002, 精确度0.000 1 g)。另外,在取样的同时,利用便携式溶氧仪(IN-SITU, AT400, 美国)对岩石底质的穴内水、砾石底质的间隙水的温度、盐度、溶氧等进行现场精确测量并记录。
1.3 数据统计分析通过物种多样性指数H' (Shannon-Weaver index) (Shannon et al, 1949)、种类丰富度d (Margalef 's index) (Margalef, 1968)、均匀度指数J' (Pielou index)(Pielou, 1968)对各站点进行统计分析,公式如下:
| $ H' = - \mathop \sum \limits_{i = 1}^S {P_i}{\text{lo}}{{\text{g}}_2}{P_i} $ | (1) |
| $ d = (S - 1)/{\text{lo}}{{\text{g}}_2}N $ | (2) |
| $ J' = H'{\text{lo}}{{\text{g}}_2}S $ | (3) |
式(1)~(3)中,S为各站点样品总数,Pi为各站点第i种的个体数与样品总个数的比值,N为各站点样品总个体数。
另外,计算各采样站点贝类的优势种类和密度,其密度为每平方米贝类的数量,优势物种比例为优势物种密度占总密度的比值。优势物种公式为:
| $ {Y_i} = {n_i}/N \times {f_i} $ | (4) |
式中,ni为各站点第i种贝类的数量,N为各站点所有种类贝类总数量,fi为某种贝类在各站点出现的频率;当某种贝类优势度Y > 0.02时,则该种贝类为该站点优势种。
2 结果 2.1 调查站点水质数据10个调查站点四季的水质数据见表 1。长山列岛底质一般为岩石或砾石;盐度随季节变化而浮动,盐度夏秋高而春冬低,平均为30.92±0.84;冬季水温最低,平均为(4.51±0.32) ℃,夏秋水温最高,平均为(21.13± 1.67) ℃;溶氧则整体表现为长山列岛主岛(南长山岛和北长山岛)调查站点较低,平均为(8.59±0.38) mg/L,而砣矶岛、大钦岛和南隍城岛调查站点溶氧相对较高,平均为(9.95±0.51) mg/L。
|
|
表 1 长山列岛10个调查站点平均水质指标 Tab.1 Average water quality index of 10 survey sites in Changshan Islands |
长山列岛不同季节同一调查站点贝类种类数量上的变化如图 2所示。4个季节采集到的贝类种类共计12种,分别是牡蛎(Crassostrea sp.)、单齿螺(Monodonta labio L.)、短滨螺(Littorina brevicula)、紫贻贝(Mytilus galloprovincialis)、矮拟帽贝(Patelloida pygmaea)、锈凹螺(Chlorostoma rustica)、疣荔枝螺(Reishia clavigera)、黄口荔枝螺(Thais luteostoma)、内饰刍秣螺(Ocinebrellus inornata)、嫁䗩(Cellana toreuma)、菲律宾蛤仔(Ruditapes philippinarum)和脉红螺(Rapana venosa)。春季采集的贝类种类最多,平均为8种;秋季次之,平均为6种;冬季最少,平均为4种。另外,南长山岛和北长山岛采集贝类种类最多,分别为9种和8种;在大钦岛采集的贝类种类最少,为6种。

|
图 2 长山列岛10个调查站点贝类种类季节变化 Fig.2 Seasonal changes of shellfish species at 10 survey sites in Changshan Islands |
通过一周年的调查发现,不同的调查点优势物种随季节的变化存在一定的变化(表 2)。牡蛎和短滨螺在四季中为优势种,紫贻贝在夏季和秋季为优势物种,矮拟帽贝在春季、秋季和冬季为优势物种,锈凹螺只在春季为优势物种,而嫁䗩在春季、夏季和冬季为优势物种。其中,春季优势物种最多,秋季次之,夏冬季最少。
|
|
表 2 不同季节贝类优势种和优势度 Tab.2 Shellfish dominant species and dominance in different seasons |
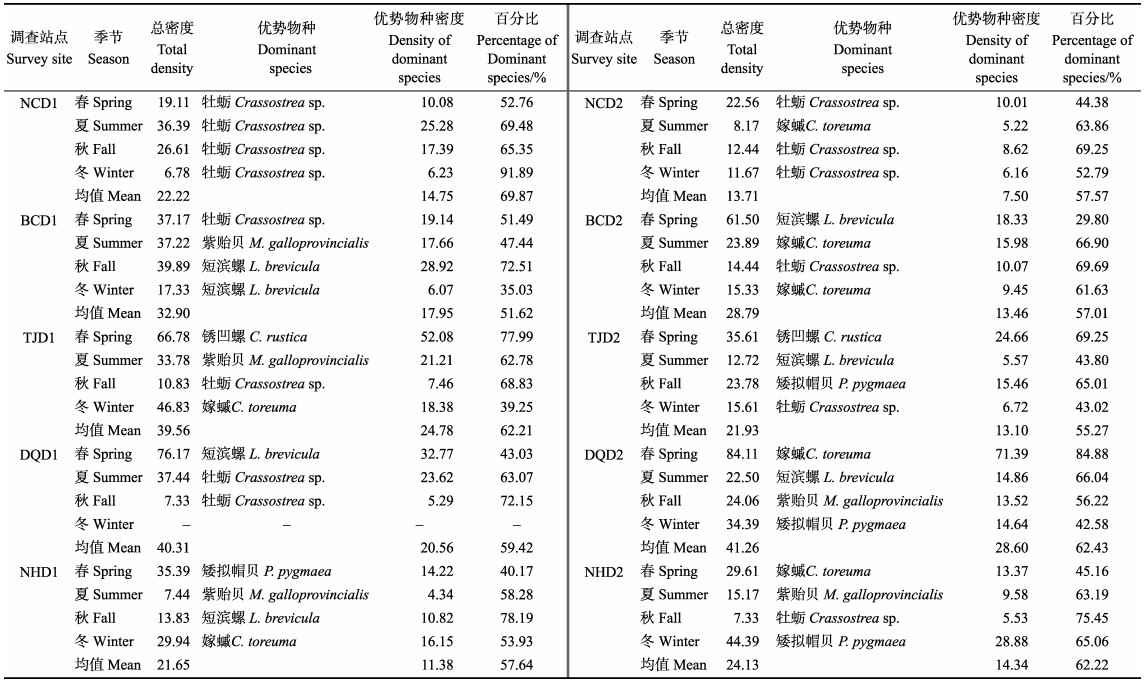
长山列岛10个调查站点潮间带贝类不同季节密度变化见表 3。不同调查站点贝类密度存在差异,其中,DQD2贝类密度最高,平均密度为41.26 ind./m2;而最低的为NCD2,平均密度为13.71 ind./m2。同一季节,不同调查站点贝类密度有所不同。其中,春季以DQD2贝类密度最高,为84.11 ind./m2;NCD1贝类密度最低,为19.11 ind./m2。夏季以DQD1贝类密最高,为37.44 ind./m2;NHD1贝类密度最低,为7.44 ind./m2。秋季以BCD1贝类密度最高,为39.89 ind./m2;NHD2和DQD1贝类密度最低,为7.33 ind./m2。而冬季以TJD1贝类密度最高,为46.83 ind./m2;最低的为NCD1,仅6.78 ind./m2。值得注意的是,同一季节,不同调查站点优势物种占比有所不同,春季以牡蛎、嫁䗩和锈凹螺密度占比较多;夏季以牡蛎、紫贻贝、短滨螺和嫁䗩占比较多;秋季以牡蛎、短滨螺和紫贻贝占比较多;而冬季以牡蛎、矮拟帽贝和嫁䗩为最多。
|
|
表 3 长山列岛10个调查站点贝类密度时空变化(单位: ind./m2) Tab.3 Temporal and spatial variation of shellfish density at 10 survey sites in Changshan Islands (unit: ind./m2) |
长山列岛10个调查站点不同季节的贝类物种多样性指数H'、均匀度指数J'和种类丰富度d统计数值见表 4。在10个调查站点中,贝类多样性指数H'平均值为1.094,其中,NCD2贝类H'为1.41,NHD2贝类H'为1.38,DQD2贝类H'为1.16,BCD2贝类H'为1.14。另外,NCD2贝类H'最高,NHD2次之,DQD1最低,为0.74。调查位点的各个潮区中贝类H'为中潮带(1.44) > 低潮带(1.40) > 高潮带(0.44)。而在10个调查站点中平均贝类H'整体上春季(1.26) > 冬季(1.21) > 秋季(1.11) > 夏季(0.95)。
|
|
表 4 10个调查站点不同季节和潮区贝类多样性指数、均匀度指数、丰富度指数 Tab.4 Shellfish diversity index, evenness index and richness index in different seasons and tidal areas of 10 survey sites |

物种均匀度指数J´在10个调查站点中平均值为0.55,其中,DQD2贝类J´为0.67,NCD2贝类J´为0.64,NHD2贝类J´为0.64,TJD1贝类J´为0.55。调查位点的各个潮区中贝类J´为中潮带(0.67) > 低潮带(0.59) > 高潮带(0.29)。各季节变化不明显。
长山列岛10个调查站点贝类丰富度平均为0.533,其中,NCD2贝类丰富度平均为0.82,NHD2贝类丰富度为0.68,在BCD2中贝类丰富度为0.60,在NCD1贝类丰富度为0.53。调查位点的各个潮区中贝类丰富度指数为中潮带(0.56) > 低潮带(0.52) > 高潮带(0.17)。而在10个调查位点中平均贝类丰富度整体上春季(0.681) > 秋季(0.573) > 夏季(0.458) > 冬季(0.418)。
3 讨论潮间带大型底栖动物空间分布与多种因素相关,包括底质(李进寿等, 2019)、游憩活动(李明峰, 2016)、潮汐(Defeo et al, 2005)和盐度(Armonies et al, 2003)等。本研究发现,长山列岛潮间带多为岩相和砾石底质,贝类多营附着或固着生活,这与李进寿等(2019)的研究结果一致。另外,本研究还观察到牡蛎、短滨螺的大量聚集,分析认为可能是由于岩相底质及干露、高温和潮汐等环境因素制约的原因,这与刘东艳等(2016)所描述的“滨螺带、藤壶–牡蛎带和海藻带”的结果一致,体现了岩相潮间带的特有现象。
有研究表明,季节变化、食物增减和水温变化是促使大型底栖动物群落结构发生变化的主要因素(Schröder et al, 2015; Mosbahi et al, 2019)。由于各潮区受到环境胁迫影响较为不同,所以生活的贝类优势物种不同。例如,高潮区为耐受性强的短滨螺,中潮区为行动能力较强的黄口荔枝螺等,以及低潮区为不耐失水的嫁䗩等。在各季节中,长山列岛各调查站点贝类种类数和密度整体表现为春、秋季节高于夏、冬季节,可能是春、秋季节食物充足、温度适宜,而夏、冬季节温度过高或过低,影响贝类对食物的获取,从而影响贝类生长(田甲申等, 2019)。夏季和冬季贝类物种丰富度较低,大多也是一些对环境耐受性强的贝类,如牡蛎和短滨螺等;春季和秋季各调查站点不仅物种丰富度高,而且密度也相对较大,如紫贻贝和矮拟帽贝等。值得注意的是,本研究发现随着季节的改变,各调查站点的优势物种也发生了一定的变化,这可能与前文所述的水温和食物的供给有关。Davison (1991)研究表明,温度是影响藻类分布和生长的重要因素,而贝类多以藻类为食,故季节变化也是温度变化,优势物种随季节的变化而改变。本研究中,矮拟帽贝在春季、秋季和冬季为优势物种,而锈凹螺只在春季为优势物种,与该观点一致。
由于潮间带的独特地理位置,容易受到人类活动的影响。研究表明,潮间带底栖生物资源量减少与开发活动和人类采捕有一定关联。李宝泉等(2020)在调查黄河三角洲潮间带后指出,潮间带生物量减少的主要原因是人类过度捕捞、开发力度大,杨颖等(2022)通过对长江口潮间带底栖生物调查也发现底栖生物资源量减少与人类采捕关系密切,这与长山列岛贝类生物量减少的原因一致。李佳等(2012)通过调查青岛沙质潮间带发现,沙滩人为活动是影响小型底栖动物数量及分布的一个因素。长山列岛旅游业发展迅速,人流密度大,对潮间带人为干扰较多,本研究认为这是影响贝类种类和生物量的主要原因。另外,大钦岛和南隍城岛养殖业的发展也是导致潮间带大型底栖生物群落结构变化的一个原因,季相星等(2018)通过对海州湾潮间带大型底栖生物调查发现,人类养殖活动对海州湾潮间带大型底栖动物群落结构产生了非常重要的影响。夏季为长山列岛的旅游旺季,人类活动量急剧增加。在长山列岛的10个调查位点中,NCD2和BCD2贝类种类最多,这可能是因为这2处调查点位于保护区内或是承包养殖海域,人类活动较少,生物量和密度最高,这与王金宝等(2018)的调查结果相吻合。NCD1和BCD1生物量和贝类种类相对较少,因为其位置处于码头和景区附近,人为干预较多。值得注意的是,DQD贝类种类最少,而DQD2贝类密度最高,本研究认为这是因为DQD2为岩石底质,且该站点是长山列岛主要的海藻尤其是海带的养殖区域,人类活动干扰多,导致短滨螺和紫贻贝等大量聚集,因而贝类种类较为单一,优势贝类种类密度较大。在北五岛中,大钦岛、南隍城岛调查位点为养殖区域,人类养殖活动频繁,故贝类种类都较为单一。砣矶岛调查位点由于处于施工附近,人类干预较为明显,因此贝类种类和数量均不多。本研究结果进一步印证了人类活动是影响长山列岛潮间带贝类生物量和物种多样性的一个重要因素。
本研究对长山列岛潮间带1周年的贝类调查共采集到贝类12种,其中优势物种有6种。王金宝等(2018)在2015—2016年对长山列岛主岛进行了四季的调查,共采集到6种贝类优势物种,包括菲律宾蛤仔、锈凹螺、紫贻贝、牡蛎、短滨螺和寇氏小节贝(Collisella kolarovai),不过本研究获取的样品并无寇氏小节贝存在,嫁䗩却是优势物种之一。本次调查结果显示,近年来长山列岛潮间带贝类优势物种略微有变化,具体变化原因还需进一步探讨。赵瑞(2014)在2012—2013年对长岛海域潮间带和潮下带进行的周年调查所获取的潮下带冬季贝类优势种为彩虹明樱蛤(Moerella iridescens),这与本研究所调查的贝类优势种也存在差异,这可能是由于本研究只局限于潮间带调查;韩庆喜等(2014)于2009—2010年对烟台3种典型潮间带(沙质底质、泥沙底质和泥质滩涂)进行调查,共采集到贝类优势物种5种,包括菲律宾蛤仔、等边浅蛤(Gomphina aequilatera)、西施舌(Coelomactra antiquata)、异白樱蛤(Macoma incongrua)和古氏滩栖螺(Batillaria cumingi)等,与本研究所获取样品存在很大不同,估计这是由二者的底质不同所致。
4 结论本研究对长山列岛10个调查站点潮间带贝类群落结构不同季节进行调查,发现不同季节贝类种类存在差异,春季采集的贝类种类最多,秋季次之,冬季最少。不同季节潮间带贝类优势物种也有所不同,表现为春季优势物种最多,秋季次之,夏冬季最少。另外,不同季节贝类密度存在时空变化,表现为春秋季节密度占比较大,夏冬季节密度占比较少。最后对长山列岛不同季节贝类物种多样性进行分析,发现在10个调查站点中,各潮区贝类物种多样性表现为中潮带(1.44) > 低潮带(1.40) > 高潮带(0.44),在季节上平均贝类物种多样性整体上表现为春季(1.26) > 冬季(1.21) > 秋季(1.11) > 夏季(0.95)。贝类物种均匀度上,不同季变化不明显。在平均贝类丰富度上,不同季节整体上为春季(0.681) > 秋季(0.573) > 夏季(0.458) > 冬季(0.418)。
近年来,长岛旅游业发展迅速,餐饮需求也日趋高档丰富,海水养殖和旅游等人类活动的增加对长山列岛海洋生态干扰日益严重。本研究开展长山列岛潮间带不同季节贝类群落结构调查,发现季节(温度)对潮间带贝类资源多样性和丰富度影响最为明显,养殖和旅游等人类活动也会对优势种群分布和数量造成影响,这些结果可为长山列岛海洋生态保护提供数据支撑。
ARMONIES W, REISE K. Empty habitat in coastal sediments for populations of macrozoobenthos. Helgoland Marine Research, 2003, 56(11): 279-287 |
BLASC J, DRAKE P, SOLE M. An integrative approach using benthos to evaluate environmental quality of the Sancti-Petri saltmarsh area (SW. Iberian peninsula). Marine Environmental Research, 2008, 66(1): 200-205 |
Compilation Committee of "China Island Chronicle". China Island Chronicle. Shandong Volume. Volume 1, the coast of northern Shandong. Beijing: Ocean Publishing House, 2013 [《中国海岛志》中国海岛志编纂委员会. 山东卷. 第1册, 山东北部沿岸. 北京: 海洋出版社, 2013]
|
DAVISON I R. Environmental effects on algal photosynthesis: Temperature. Journal of Phycology, 1991, 27(1): 2-8 DOI:10.1111/j.0022-3646.1991.00002.x |
DEFEO O, MCLACHLAN A. Patterns, processes and regulatory mechanisms in sandy beach macrofauna: A multi-scale analysis. Marine Ecology Progress, 2005, 295(2): 1-20 |
HAN Q X, YUAN Z Y, CHEN B J, et al. The community structure and distribution pattern of intertidal macrobenthos in the intertidal zone of Yantai. Marine Sciences, 2014, 38(9): 59-68 [韩庆喜, 袁泽轶, 陈丙见, 等. 烟台潮间带大型底栖动物群落组成和结构研究. 海洋科学, 2014, 38(9): 59-68] |
JI X X, LI J, JIANG Y, et al. Structure characteristics and seasonal variations of macrobenthic communities in the intertidal zone of Haizhou Bay from 2012 to 2017. Environmental Monitoring in China, 2018, 34(6): 130-136 [季相星, 李军, 姜毅, 等. 2012—2017年海州湾潮间带大型底栖动物群落结构及变化. 中国环境监测, 2018, 34(6): 130-136] |
JIN W Y, PENG X, WANG N, et al. Study on the macrobenthic community structure in the intertidal zone of the Yushan Islands in summer. Marine Science, 2017, 41(3): 17-25 [金文育, 彭欣, 王宁, 等. 渔山列岛夏季潮间带大型底栖生物群落结构研究. 海洋科学, 2017, 41(3): 17-25] |
LI B Q, JIANG S Y, LÜ J Z, et al. Species composition and long-term variation of macrobenthos in intertidal zone and offshore areas of the Yellow River Delta. Biodiversity Science, 2020, 28(12): 1511-1522 [李宝泉, 姜少玉, 吕卷章, 等. 黄河三角洲潮间带及近岸浅海大型底栖动物物种组成及长周期变化. 生物多样性, 2020, 28(12): 1511-1522] |
LI J S, RUAN J F, LUO F. Benthic community and ecological characteristics in the rock intertidal zone of Xiahu shore. Journal of Ningde Normal University (Natural Science), 2019, 31(4): 390-394 [李进寿, 阮俊峰, 罗芬. 下浒海滨岩相潮间带底栖动物群落及其生态特征. 宁德师范学院学报(自然科学版), 2019, 31(4): 390-394] |
LI J, HUA E, ZHANG Z N. Distribution and seasonal dynamics of meiofauna in intertidal zone of Qingdao sandy beaches, Shandong Province of East China. Chinese Journal of Applied Ecology, 2012, 23(12): 3458-3466 [李佳, 华尔, 张志南. 青岛砂质潮间带小型底栖动物分布及季节动态. 应用生态学报, 2012, 23(12): 3458-3466] |
LI M F. Macrobenthos community distribution in intertidal beach and its response to recreational activities. Journal of Central South University of Forestry and Technology, 2016, 36(12): 102-107 [李明峰. 沙滩潮间带大型底栖动物群落分布及其对游憩活动的响应. 中南林业科技大学学报, 2016, 36(12): 102-107] |
LIN H S, WANG J J, HUANG Y Q, et al. An ecological study on the benthos in intertidal zone of Xinghua Bay. Progress in Fishery Sciences, 2016, 37(1): 15-21 [林和山, 王建军, 黄雅琴, 等. 兴化湾潮间带大型底栖生物群落生态研究. 渔业科学进展, 2016, 37(1): 15-21] |
LIU D Y, HAN Q Y. Intertidal zone survey method and practice. Beijing: Science Press, 2016 [刘东艳, 韩秋影. 潮间带调查方法与实践. 北京: 科学出版社, 2016]
|
LIU S L, QIN X D, WANG G J, et al. Structure and diversity of macrozoobenthic communities in intertidal zone with mangrove forest in Fengjiajiang Estuary, Beihai City in summer in 2017. Wetland Science, 2019, 17(3): 352-358 [刘士龙, 秦旭东, 王广军, 等. 2017年夏季北海市冯家江入海口红树林潮间带大型底栖动物群落结构及多样性. 湿地科学, 2019, 17(3): 352-358] |
MARGALEF R. Perspectives in ecological theory. Chicago: University of Chicago Press, 1968: 1-111
|
MENG S, FANG J H, JIANG Z J, et al. Experimental study of the choice behavior of Perinereis aibuhitensis Grube among different sediment types. Progress in Fishery Sciences, 2020, 41(4): 110-116 [孟珊, 房景辉, 蒋增杰, 等. 双齿围沙蚕对潮间带不同类型底质选择行为的研究. 渔业科学进展, 2020, 41(4): 110-116] |
MOSBAHI N, SERBAJI M M, PEZY J P, et al. Response of benthic macrofauna to multiple anthropogenic pressures in the shallow coastal zone south of Sfax (Tunisia, central Mediterranean Sea). Environmental Pollution, 2019, 253(1): 474-487 |
NIU L H, ZOU G H, GUO Y T, et al. Eutrophication dangers the ecological status of coastal wetlands: A quantitative assessment by composite microbial index of biotic integrity. Science of the Total Environment, 2022, 816(6): 151620 |
PIELOU E C. Ecological diversity. New York: John Wiley, 1975, 1-163 |
SCHRÖDER M, SONDERMANN M, SURES B, et al. Effects of salinity gradients on benthic invertebrate and diatom communities in a German lowland river. Ecological Indicators, 2015, 57(1): 236-248 |
SHANNON C E, WEAVER W. The mathematical theory of communication. Urbana: University of Illinois Press, 1949: 1-144
|
SHI Q. Spatiao-temporal modes and circulation variation on the interannual variation of seasonal mean wind-driven current field in the Bohai Sea and Yellow Sea in winter and summer. Journal of Applied Oceanography, 2019, 38(1): 93-108 [石强. 渤、黄海冬、夏季节风生流场年际变化时空模态与环流变异. 应用海洋学学报, 2019, 38(1): 93-108] |
TIAN J S, WANG Z, LI D H, et al. Effects of temperature, and diet on stable carbon and nitrogen isotopes discrimination in different species of filtering mollusks. Journal of Dalian Ocean University, 2019, 34(1): 80-86 [田甲申, 王震, 李多慧, 等. 温度、饵料和物种对滤食性贝类碳氮稳定同位素分馏的影响. 大连海洋大学学报, 2019, 34(1): 80-86] |
WANG J B, LI X Z, WANG H F, et al. Macrobenthic ecology in the interridal zones of Changdao Islands in Shandong Province. Ocean Science, 2018, 42(10): 41-52 [王金宝, 李新正, 王洪法, 等. 山东长岛潮间带大型底栖动物生态特征研究. 海洋科学, 2018, 42(10): 41-52] |
WU Q E, SUN D R, LI C H. The characteristics of macrobenthic community in intertidal zone of Nanpeng Islands, Guangdong Province. Progress in Fishery Sciences, 2010, 31(4): 101-106 [吴洽儿, 孙典荣, 李纯厚. 广东南澎列岛潮间带大型底栖生物的群落特征. 渔业科学进展, 2010, 31(4): 101-106] |
YANG Y, CHEN S S, ZHOU H H, et al. Ecological evaluation and variation trend analysis of macrobenthos in intertidal zone of Changjiang Estuary. Acta Ecologica Sinica, 2022, 42(4): 1606-1618 [杨颖, 陈思思, 周红宏, 等. 长江口潮间带底栖生物生态及变化趋势. 生态学报, 2022, 42(4): 1606-1618] |
YU F, ZHANG Z X, DIAO X Y, et al. Analysis of evolution of the Huanghai Sea cold water mass and its relationship with adjacent water masses. Acta Oceanologica Sinica, 2006(5): 26-34 [于非, 张志欣, 刁新源, 等. 黄海冷水团演变过程及其与邻近水团关系的分析. 海洋学报(中文版), 2006(5): 26-34] |
ZHAO R. The ecological study of macrobenthos in the waters around Changdao Islands. Master´s Thesis of Ocean University of China, 2014 [赵瑞. 长岛海域大型底栖动物生态学研究. 中国海洋大学硕士研究生学位论文, 2014]
|
ZHENG D, ZHANG R A. Analysis of spring water mass off the coast of Yanwei and Shidao. Marine Bulletin, 1983(1): 61-68 [郑东, 张瑞安. 烟威及石岛近海春季水团分析. 海洋通报, 1983(1): 61-68] |