



2. 山东长岛近海渔业资源国家野外科学观测研究站 山东 烟台 265800
2. National Field Observation and Research Center for Fisheries Resources in Changdao Waters, Yantai 265800, China
渤海湾为三面陆缘环抱、东部与渤海毗连的半封闭性海湾,地处暖温带北缘、季风气候区,在陆上气候和黄海冷水团次级水团–渤海水团交互影响下,温、盐等季节变化显著,同时,该海域受渤海逆时针环流、往复流性质的潮流及M2潮汐余流影响,形成错综复杂的海洋生态环境;来自黄海北部的外海高盐水系和渤南沿岸低盐水系(前者夏季为低温,冬季为高温;后者夏季为高温,冬季为低温)的分布、演变和消长过程构成了渤海湾基本水文特征(农业部水产局和农业部黄海区渔业指挥部, 1990)。渤海湾沿岸有滦河、蓟运河、潮白新河、海河、独流减河、子牙新河、漳卫新河和徒骇河等河流入海带来的淡水和营养盐类,促进了饵料生物大量繁殖,为渔业生物提供了适宜的生长环境,历史上为多种鱼类(夏世福, 1960; 吴光宗等, 1983; 杨东莱等, 1984; 姜言伟等, 1988; 孔立波等, 1996)、虾类(邓景耀, 1960、1980)、蟹类(邓景耀等, 1988)和口足类(邓景耀等, 1988; 谷德贤等, 2018)等的产卵场及其幼体的主要索饵场,同时形成了渤海三大渔场之一——渤海湾渔场。
渤海湾作为一个浅滩淤泥质海湾,海底地形自岸向海倾斜,沿岸受人类活动影响较大,半封闭的港湾特征决定其与外海交换能力差(赵维等, 2019)。近40年来,渤海湾周围集中了海洋化工、海上石油开采、港口、滩涂养殖和盐业等多种经济活动(聂红涛等, 2008);大规模围垦滩涂、临海城市和港口码头建设导致岸线人工化程度逐年增高(至2015年渤海湾沿岸人工岸线比例约占岸线总长度的95%以上) (孙百顺等, 2017)及潮流场变化(孙晓宇等, 2014),湾内环流涡旋增多(王勇智等, 2015; 白玉川等, 2021),水交换率下降(赵维等, 2019);大量陆源污染物质排入渤海湾,其一直处于富营养化状态,赤潮频发(Wang et al, 2009; 赵海萍等, 2016; 刘晶晶等, 2021),并叠加镉、汞、铅和石油烃复合污染更加剧了渤海湾生态负担(许思思等, 2010)。在全球变化背景下,渤海湾海表温持续上升,海洋热浪发生频次、持续时间和累积强度显著增加(自然资源部国家海洋信息中心,2019—2022);黄河和海河等入海径流量下降、黄河尾闾摆动和海河水系诸河口建闸(雷坤等, 2007; Wu et al, 2020)等使渤海湾淡水和营养盐输入通量显著下降(雷坤等, 2007),温盐呈显著上升态势(吴德星等, 2004),严重影响河口生态功能和渔业生物饵料供给,为鱼类产卵场环境生态带来严重负面影响。特别是1960s以来,渤海湾渔业资源遭遇了生长型(冯志青等, 1982、1987)和补充型(农业部水产局和农业部黄海区渔业指挥部, 1991)的双重捕捞过度,资源呈现严重衰退(程济生等, 2004)。
渔业种群的变动主要由补充量变化驱动,产卵场能为鱼类早期资源(鱼卵和仔稚幼鱼)提供生长、发育和存活的基本条件,鱼类早期资源的存活率则决定了种群世代强弱((Houde, 1987)。鱼卵、仔稚鱼调查提供了一种只需浮游生物网就可对产卵场中大部分鱼种丰度进行取样和评估的可能性(Sherman et al, 1984; Koslow et al, 2016),是鱼类繁殖生物学和补充机制研究的有效方法(Sassa et al, 2019)。特别对那些不便直接开展资源调查的鱼类,相较传统底拖网调查优势更加显著(Saville et al, 1981);调查获取的鱼类早期资源量及产卵场面积和位置等信息可帮助确定产卵亲体种类、产卵期、种群特别是复合种群(meta-population)的划分(Hare et al, 2013),并可直接作为种群补充成功率和亲体量评估的重要指标(Sassa et al, 2019);通过高质量、长周期鱼卵仔稚鱼调查数据能获取非捕捞因素导致的产卵亲体量和产卵场年际变异信息(Lasker, 1985; Hsieh et al, 2005; 2006; Lo et al, 2016; Takasuka et al, 2019; Nielsen et al, 2020; Thompson et al, 2021),其为研究人类活动(Hsieh et al, 2005、2006)和气候变化(Nielsen et al, 2021; Thompson et al, 2021)对海洋鱼类种群影响的重要信息。渤海湾水域也为作者所在单位鱼类早期资源调查资料积累较丰富的海区之一,1982—2019年,中国水产科学研究院黄海水产研究所先后在渤海湾海域开展过60余航次鱼卵、仔稚鱼调查(姜言伟等, 1988; 万瑞景等, 1998; 程济生等, 2004; 卞晓东等, 2018),最近又于2020年4—12月间开展了周年逐月调查,取得了长期连续观测资料。为系统掌握渤海湾鱼类早期资源群落结构特征及其长期演替过程,本研究选取其中53个季度月航次数据来进行综合分析,以期为渤海湾鱼类早期资源评价与保护研究积累基础资料。
1 材料与方法 1.1 数据采集和样品分析选取涵盖渤海湾1980s、1990s、2000s—2010s前期、2010s中期、2010s晚期和2020s前期(含年内各季节和鱼类主要产卵期) 53航次数据,研究各调查时期鱼类早期资源种类组成、资源丰度、优势种类和物种多样性水平,产卵和育幼场范围及重心迁移,并对产卵亲体产卵、适温和栖息类型开展分析,各调查时期采样站位及时间详见图 1和表 1。

|
图 1 不同调查时期渤海湾鱼卵、仔稚鱼采样区域和调查站位
Fig.1 Survey area and sampling stations to fish eggs and larvae in the Bohai Bay in different survey periods
a和d:1980s站位(a. 1982年5、6、8、10月;d. 1983年1—2月);b和e:1990s站位(b. 1992年8、10月,1993年5—6月,1998年5—6、8、10月;e. 1993、1999年2月);c和f:2000—pre2010s站位(c. 2004年5月,2012年6、8月,2009年10月,2013、2014年5、6、8、10月;f. 2014年2月,2015年1月);g和j:mid2010s站位(g. 2015、2016年5、6、8月,2016年10月;j. 2016年1、12月);h和k:late2010s站位(h. 2017、2018年5、6、8、10月,‘ ’所示站位仅在2017年5、6月开展;k. 2017年3、11—12月);i和l:pre2020s站位(i. 2020年5、6、8、10月;l. 2020年12月)。
a and d: Sampling stations in the1980s (a. May, June, August and October in 1982; d. January–February in 1983); b and e: Sampling stations in the1990s (b. August, October in 1992, May–June in 1993, May–June, August and October in 1998; e. February in 1993 and 1999); c and f: Sampling stations in the 2000s to pre2010s (c. May in 2004, June and August in 2012, October in 2009, May, June, August and October in 2013 and 2014; f. February in 2014 and January in 2015); g and j: Sampling stations in the middle 2010s (g. May, June, and August in 2015 and 2016, October in 2016; j. January and December in 2016); h and k: Sampling stations in the late 2010s (h. May, June, August and October in 2017 and 2018, stations marked as '' only surveyed in May and June of 2017; k. March, November–December in 2017); i and l: Sampling stations in the early 2020s (i. May, June, August and October in 2020; l. December in 2020). ’所示站位仅在2017年5、6月开展;k. 2017年3、11—12月);i和l:pre2020s站位(i. 2020年5、6、8、10月;l. 2020年12月)。
a and d: Sampling stations in the1980s (a. May, June, August and October in 1982; d. January–February in 1983); b and e: Sampling stations in the1990s (b. August, October in 1992, May–June in 1993, May–June, August and October in 1998; e. February in 1993 and 1999); c and f: Sampling stations in the 2000s to pre2010s (c. May in 2004, June and August in 2012, October in 2009, May, June, August and October in 2013 and 2014; f. February in 2014 and January in 2015); g and j: Sampling stations in the middle 2010s (g. May, June, and August in 2015 and 2016, October in 2016; j. January and December in 2016); h and k: Sampling stations in the late 2010s (h. May, June, August and October in 2017 and 2018, stations marked as '' only surveyed in May and June of 2017; k. March, November–December in 2017); i and l: Sampling stations in the early 2020s (i. May, June, August and October in 2020; l. December in 2020).
|
|
|
表 1 各调查时期渤海湾鱼卵、仔稚鱼调查时间和站位数 Tab.1 Survey time and number of sampling stations to fish eggs and larvae during different survey periods in the Bohai Bay |
各调查时期鱼卵、仔稚鱼样品采集均采用网口内径80 cm、长270 cm、36 GG (孔径0.505 mm)筛绢制成的大型浮游生物网,采用水平拖网,每站表层拖网10 min,拖曳速度2 nmile/h。标本用5%海水甲醛溶液固定。实验室内从大型浮游生物网采集样品中挑出鱼卵和仔稚鱼。样品全样本分析,体视显微镜下经形态学鉴定并反复核实,确认至每一物种单元,并按种类及其发育阶段分别记录个体数。发育阶段包括卵(分裂期、原肠期、胚胎期、孵化期和死卵)和仔稚幼鱼(前期仔鱼、后期仔鱼、稚鱼和幼鱼)。鱼种学名参照World Register of Marine Species (WoRMS, https://www.marinespecies.org/)。
1.2 鱼卵仔稚鱼生态密度和种类数鱼卵、仔稚鱼生态密度为个体数生态密度(ecological density of number in early life history, EDN-ELH)。不同调查时期鱼卵EDN-ELH取渤海湾硬骨鱼类主要产卵季节(5、6、8和10月)出现的全部鱼卵数据来计算;仔稚鱼EDN-ELH取周年各月调查(5、6、8、10、11月至次年3月)出现的全部仔稚鱼数据来计算;在数据整理过程中,由于水平拖网速度难以严格控制,为便于同质比较和数据质量控制,样品定量分析时以每站每网实际捕获鱼卵、仔稚鱼(ind./haul)作为指标进行比较(姜言伟等, 1988; 万瑞景等, 1998; 卞晓东等, 2018、2022a、b)。因不同调查时期各月调查站位和数量不同,以1982—1983年调查结果作为本底(Yref, 年效应值为1),采用R中广义线性模型(generalized linear model, GLM),考虑捕捞努力量的空间分布(NR, 渔区zone)和数据尺度特征(NS, 调查时间month),分别对各调查时期(Y, 年份year)鱼卵或仔稚鱼EDN-ELH (CPUEi)进行标准化,构建资源丰度指数(abundance index, AI)指示不同调查时期鱼卵、仔稚鱼和各种EDN-ELH (卞晓东等, 2018、2022a、b)。计算公式参照官文江等(2014)和官文江(2015):
| $ {\text{CPUE}} = \frac{{qaN}}{A} = aqD $ | (1) |
式(1)为CPUE与资源密度(EDN-ELH)一般表达式。式中,q为捕捞系数,N为资源量,A为渔场面积,D为平均EDN-ELH,a为有效系数,即渔具能有效覆盖或能接近的资源比例。
| $ E({\text{CPU}}{{\text{E}}_{i, j, k, e, g}}) = {H_h}{G_g}{Y_i}{Q_j}{R_k}{a_0}{q_0}{D_0} $ | (2) |
式(2)为CPUE与a、q及N或D的函数关系。式中,a0为有效系数,q0标准捕捞系数,D0为标准时间–区域下的EDN-ELH,Hh为环境h的相对效应(相对于标准环境, 下同),Gg为渔具g的相对效应,Yi为年i的相对效应,Qj为月j的相对效应,Rk为区域k的相对效应。
| $ {\text{CPU}}{{\text{E}}_i} = \frac{1}{{{N_{\text{S}}}}}\sum\limits_{j = 1}^{{N_{\text{S}}}} {\sum\limits_{k = 1}^{{N_{\text{R}}}} {{A_k}E({\text{CPU}}{{\text{E}}_{i, j, k, 0, 0}})} } $ | (3) |
式(3)为去除a与q的变化后CPUE与资源量或EDN-ELH的正比关系,即标准化的CPUE。式中,NS为调查月份,NR为渔区数,CPUEi, j, k, 0, 0为忽略环境效应与渔具效应下(即假设Hh与Gg为1)的CPUE。将式(2)带入式(3)可得式(4):
| $ {\text{CPU}}{{\text{E}}_i} = {a_0}{q_0}{D_0}{Y_i}\left[ {\frac{1}{{{N_{\text{S}}}}}\sum\limits_{j = 1}^{{N_{\text{S}}}} {\sum\limits_{k = 1}^{{N_{\text{R}}}} {{A_k}{Q_j}{R_k}} } } \right] $ | (4) |
由于CPUE仅代表相对资源丰度,CPUE时间系列除以相同的数不会改变资源量的相对关系。当各年CPUE均除以参考年份的CPUE,则获得式(5)相对丰度指数Ii (官文江等, 2014; 官文江, 2015):
| $ {I_i} = \frac{{{\text{CPU}}{{\text{E}}_i}}}{{{\text{CPU}}{{\text{E}}_{{\text{ref}}}}}} $ | (5) |
将式(4)代入式(5),则得式(6):
| $ {I_i} = \frac{{{Y_i}}}{{{Y_{{\text{ref}}}}}} $ | (6) |
式(5)和式(6)中,Ii为相对年效应,CPUEi为第i年CPUE,CPUEref为参考年份的CPUE。
鱼卵或仔稚鱼种数为不同调查时期各航次出现的鱼卵或仔稚鱼物种数。
1.3 鱼卵、仔稚鱼的分布及重心迁移根据各调查时期(表 1)实际调查获取渤海湾鱼类早期资源(鱼卵或仔稚鱼)EDN-ELH,通过ArcGIS10.4软件地理统计分析模块的Empirical Bayesian Kriging插值处理后,绘制不同调查时期产卵(育幼)场分布图。
采用Garrison分布重心法(张雨轩等, 2022a)计算各调查时期(年份)鱼卵(仔稚鱼)分布重心,计算公式为:
| $ X=\frac{{\displaystyle \sum _{i=1}^{n}({X}_{i}\times {\rho }_{i})}}{{\displaystyle \sum _{i=1}^{n}{\rho }_{i}}} $ | (7) |
| $ Y=\frac{{\displaystyle \sum _{i=1}^{n}({Y}_{i}\times {\rho }_{i})}}{{\displaystyle \sum _{i=1}^{n}{\rho }_{i}}} $ | (8) |
式(7)和式(8)中的X、Y分别为产卵(育幼)重心的经、纬度,Xi、Yi分别为i站位的经、纬度,
运用Pinkas相对重要性指数[index of relative importance, IRI, 公式(9)](Pinkas et al, 1971)对每航次调查出现鱼卵和仔稚鱼种类组成分别进行分析,以确定鱼卵或仔稚鱼优势种类(dominant species)、重要种类(important species)和主要种类(main species)的成分:
| $ \text { IRI }=(N \%+W \%) \times F \%$ | (9) |
| $ \mathrm{IRI}=N \% \times F \%$ | (10) |
式(9)中,N%为每航次调查过程中某种鱼卵或仔稚鱼个体数量占鱼卵或仔稚鱼总量比例;W%为每航次调查过程中某种鱼卵或仔稚鱼生物量比例。由于鱼卵和仔稚幼鱼个体都很小,因此不考虑生物量,只考虑个体数量这一因素,IRI的计算公式可以简化为公式(10) (卞晓东等, 2018、2022a、b)。取IRI>1 000的种类定义为鱼卵或仔稚鱼优势种;鱼卵重要种类IRI值为200~1 000;仔稚鱼重要种类IRI值为100~1 000。
1.5 鱼卵仔稚鱼物种多样性水平及其种类更替采用α多样性测度方法,香农–威纳指数[Shannon- Wiener Index, H′公式(11)]和辛普森多样性指数[Simpson's Diversity Index, DS公式(12)]研究渤海湾不同调查时期各季节鱼卵和仔稚鱼物种多样性;采用β多样性测度方法,Jaccard群落种类组成相似性指数[coefficient of community, CC公式(13) (Whittaker, 1972)]和基于鱼类早期资源种类相似程度的层次聚类方法比较各年间鱼类早期资源(鱼卵仔稚鱼种数合并计数)种类相似程度(卞晓东等, 2018、2022b)。
公式(11)和(12)中,S为不同调查时期各季节鱼类早期资源中出现的鱼卵或仔稚鱼种类数,Pi为群落中第i种鱼卵或仔稚鱼所占鱼卵或仔稚鱼总量的个体比例。公式(13)中,Ss为2个比较年份间鱼类早期资源共有种类数,Sj和Sk为2个比较年份各自拥有种数。当CC为0~0.25时,群落之间极不相似;当CC为0.25~0.50时,群落之间中等不相似;当CC为0.50~0.75时;群落之间中等相似;当CC为0.75~1.00时,群落之间极相似(Whittaker, 1972; 卞晓东等, 2018)。
| $ H' = - \mathop \sum \limits_{i = 1}^{\text{s}} {P_i}\ln {P_i} $ | (11) |
| $ {D_S} = 1 - \mathop \sum \limits_{i = 1}^{\text{s}} P_i^2 $ | (12) |
| $ CC = S_{s}/(S_{j} + S_{k} –S_{s})$ | (13) |
渤海湾鳀(Engraulis japonicas)、青鳞小沙丁鱼(Sardinella zunasi)、斑鰶(Konosirus punctatus)、赤鼻棱鳀(Thryssa kammalensis)、黄鲫(Setipinna tenuifilis)、蓝点马鲛(Scomberomorus niphonius)和花鲈(Lateolabrax maculatus)等鱼卵EDN-ELH取主要产卵季节(5、6、8和10月)数据来计算,各种AI计算方法同1.2中鱼卵、仔稚鱼AI计算方法。运用Origin 2018软件绘制1980s以来渤海湾主要鱼类鱼卵资源丰度指数变动图。
1.7 亲体产卵、适温和栖息类型分析依据卵子类型将亲体划分为产浮性卵鱼类(pelagic egg, P)、产黏着沉性卵鱼类(demersal egg, D)、产附着性卵鱼类(egg with adhesive filaments, AF)、产凝集浮性卵鱼类(agglutinative pelagic egg, AP)和卵胎生鱼类(ovoviviparity, O)。根据鱼类区系分类有关文献索引(《中国渔业资源调查和区划》编辑委员会, 1990; 田明诚等, 1993)将渤海湾亲体适温类型划分为暖温种(warm temperate species, WT)、暖水种(warm water species, WW)和冷温种(cold temperate species, CT)。根据刘静等(2011)鱼类栖息类型划分将渤海湾亲体划分为大陆架浅水底层鱼类(continental shelf demersal fish, CD)、大陆架浅水中底层鱼类(continental shelf benthopelagic fish, CBD)、大陆架浅水中上层鱼类(continental shelf pelagic-neritic fish, CPN)、大陆架岩礁性鱼类(continental shelf reef-associated fish, CRA)、大陆架大洋洄游性中上层鱼类(oceanic pelagic fish, OEP)和大洋深水底层鱼类(oceanic bathydemersal fish, OMP)。
2 结果与分析 2.1 渤海湾鱼卵总种类数和生态密度季节变化和年代际变化近40年来,渤海湾(5、6、8和10月)鱼卵种数由1980s的27种,1990s前期的20种、后期的19种,逐次下降至2000s—2010s初期的11种(2014—2015年,不足1980s的50%);自2010s中期(2015—2016年)开始,鱼卵种数逐步回升,至2018—2019年达16种,最近2020—2021年为22种(图 2a)。季节变化上,各调查时期春季(5月)和春夏季(6月)采集浮性卵种数最多,夏季(8月)次之,秋季(10月)种类较少,冬季(11月至次年3月)均未采集到浮性卵(图 3a)。长期变化上,春季、春夏季和夏季采集鱼卵种数年代际波动明显,呈先下降后上升的变动趋势。春季鱼卵种数由1980s的19种,1990s初期的18种、后期的15种,逐次下降至2000s—2010s初期的4种(不足1980s的25%);此后,鱼卵种数逐步回升,至2020—2021年恢复至17种(图 3a)。春夏季,鱼卵种类由1980s的15种,1990s初期的18种、后期的15种,降至2000s—2010s初期的7种(2004—2015年各年均不足1980s的50%);自2010s中后期开始,鱼卵种数逐步增多,2018—2019年达18种;至2020—2021年为13种(图 3a)。夏季,鱼卵种类由1980s的10种,1990s初期的8种、后期的6种,逐渐降至2000s—2010s初期的3~4种(2013—2014、2014—2015年各3种,均不足1980s的50%);自2010s中期开始,鱼卵种数逐步回升,至2018—2019年为8种,当前2020—2021年为6种。秋季出现鱼卵种数相对稳定,在1~3种之间(图 3a)。

|
图 2 渤海湾各调查时期(年份)鱼卵(a)仔稚鱼(b)资源丰度指数和种类数 Fig.2 Abundance index and species number of (a) fish eggs and (b) fish larvae across different survey periods (year) since the 1980s in the Bohai Bay |

|
图 3 渤海湾各调查时期(年份)不同季节鱼卵、仔稚鱼种类数和生态密度概况 Fig.3 Number of species and ecological density to fish eggs and larvae profiles along the survey season across different survey periods in the Bohai Bay a:鱼卵种类;b:鱼卵生态密度;c:仔稚鱼种类;d:仔稚鱼生态密度 a: Number of egg species; b: Ecological density of fish eggs; c: Number of larvae species; d: Ecological density of fish larvae |
如果1982—1983年鱼卵AI为1,那么此后其资源丰度指数变动为0.21~2.09倍,历史低值在2014—2015年,历史高值在1992—1993年。鱼卵AI呈先上升后下降而后又上升的变动趋势,由1980s的1.00跃升至1990s初期的2.09,此后AI急剧下跌,至1990s后期为0.24;2000s—2010s前期,AI一直处于低水平区间(0.21~0.26),在2014—2015年跌至历史低值0.21;自2010s中期开始,AI逐步回升,特别是2017年后,AI恢复趋势明显,2018—2019年升至历史次高值1.61,最近2020—2021年AI为0.88。季节变化上,鱼卵平均资源密度(AEDN, ind./haul)在各调查时期升温季节(春季和春夏季)较高;夏季鱼卵AEDN急剧下降,至秋季整个调查海区仅有零星鱼卵分布(图 3b);冬季无浮性鱼卵分布。长期变化上,不同年份春季AEDN差异极显著(Kruskal-Wallis H-Test for one-way ANOVA by ranks, H(10, n=204)=81.97,P<0.001;各年均值比较χ2=59.68, df=10, P<0.001),由1980s的4 869.5 ind./haul升至1990s初期的历史高值30 666.0 ind./haul,1990s后期急剧跌至292.2 ind./haul,在2004—2015年AEDN徘徊在历史低值区间16.5~542.3 ind./haul (2013—2014年跌至历史低值16.5 ind./haul,不足1980s的0.4%);自2010s中后期开始,鱼卵AEDN呈波动上行趋势,至2018—2019年为3 718.9 ind./haul,2020—2021年则高达25 159.5 ind./haul (图 3b)。春夏季鱼卵AEDN波动趋势与春季类似,不同年份AEDN差异极显著(Kruskal-Wallis H-Test for one-way ANOVA by Ranks,H(10, n=231)=68.92,P<0.001;各年均值比较χ2=55.54,df=10,P<0.001),1980s为70.61 ind./haul,1990s初跃升至30 666.0 ind./haul;至1990s后期迅速降至292.6 ind./haul,2000s—2010s初期徘徊在历史低值区间27.9~98.55 ind./haul (2013—2014年至历史低值27.9 ind./haul);自2010s中期开始,AEDN呈波动上行趋势,且2010s后期始,AEDN均在1 000 ind./haul之上,至2020—2021年为1 130.4 ind./haul (图 3b)。长期变化上,不同调查年份夏季鱼卵AEDN间差异不及春季和春夏季显著(Kruskal-Wallis H-Test for one-way ANOVA by ranks,H(10, n=213)=21.49,P<0.05;各年均值比较χ2=18.01,df=10,P=0.055>0.05),夏季AEDN自1982—2015年间呈下降趋势,由1980s的238.9 ind./haul降至2014—2015年历史低值1.54 ind./haul;自2010s中后期开始,AEDN波动上行,于2015—2016年达历史高值263.2 ind./haul。长期变化上,不同调查年份秋季AEDN差异显著(Kruskal-Wallis H-Test for one-way ANOVA by ranks,H(9, n=175)=81.34,P<0.001;各年均值比较χ2=71.17,df=9,P<0.001)。秋季鱼卵AEDN自1980s历史高值100.9 ind./haul降至1990s前期的10.6 ind./haul,此后AEDN一直在0.28~4.64 ind./haul较低水平区间内波动,至2020—2021年仅为0.28 ind./haul,资源衰退严重(图 3b)。
2.2 渤海湾仔稚鱼种类数和生态密度季节变化和年代际变化近40年来,渤海湾周年调查仔稚鱼种数由1980s的27种、1990s前期的25种、后期的22种,逐次下降至2000s—2010s初期的10~15种低位区间(2004—2012年仅10种,约为1980s的37%);2010s中后期仔稚鱼种数回升,2017—2018年升至28种,2020—2021年为26种(图 2b)。季节变化,各调查时期仔稚鱼种数在升温季呈上升趋势,并在春夏季至年内高值;夏季仔稚鱼种数下降,秋季出现种数进一步下降,冬季至年内最低(个别调查年份除外)但仍有一定种类仔稚鱼出现(图 3c)。长期变化上,春季、春夏季、夏季和秋季出现仔稚鱼种数均主要呈先下降后上升的变动趋势。春季出现仔稚鱼种数由1980s的15种逐次降至2000—2010s前期的3~7种,2014—2015年至历史低值3种;2010s中后期升至6~15种(图 3c),当前2020—2021年为12种。春夏季出现仔稚鱼种数自1980s—1990s前期小幅上升,由13种升至15种,此后种类数逐渐降低,在2000s—2010s前期剧降至5~6种,2014—2015年至历史低值5种;此后,2010s中期始仔稚鱼种数逐步升至10~16种,2017—2018年至历史高值16种,2020—2021年为13种(图 3c)。夏季出现仔稚鱼种数由1980s年的10种升至1990s前期的15种、1990后期14种,此后在2000—2010s前期剧降至3~5种,2014—2015年至历史低值3种;2010s中期开始,种数上升至8~11种,在2018—2019年至历史高值11种,2020—2021年为9种(图 3c)。秋季出现仔稚鱼种数也由1980s和1990s前期的5种降至1990s末期的1种,2000s—2010s前期至0~1种低水平区间,2013—2014年至历史低值0种;2010s中后期开始,仔稚鱼种数逐年递增,在2020—2021年恢复至5种(图 3c)。冬季出现仔稚鱼种数较少,长期变化呈明显上升趋势。1980s和1990s仅出现1~2种,2000s—2010s前期升至2~4种;2010s中后期至1~4种,最近2020—2021年至历史高值6种(图 3c)。
如果1982—1983年仔稚鱼AI为1.00,那么此后其AI变动为0.25~1.33倍,历史低值在2004—2012年,历史高值在2020—2021年。仔稚鱼AI呈先下降后上升的变动趋势,AI值由1980s和1990s初期的1.00降至2004—2012年的0.25,此后AI波动上升,至2017—2018年恢复至1980s水平,近年来,仔稚鱼AI值在0.35~1.33区间剧烈波动,最近2020—2021年为1.33。季节变化,仔稚鱼AEDN在各调查时期升温季节较高,并在春夏季升至年内最高值;夏季仔稚鱼AEDN急剧下降,至秋季仔稚鱼AEDN降至年内最低,冬季仍有一定数量的仔稚鱼分布(图 3d)。长期变化上,相同季节仔稚鱼AEDN均呈剧烈年际和年代际波动(图 3d)。不同调查年份,春季AEDN差异极显著(Kruskal-Wallis H-Test for one-way ANOVA by Ranks,H(10, n=204)=81.97,P<0.001;各年均值比较χ2= 59.68,df=10,P<0.001),呈先上升后下降而后又波动上升的趋势,由1980s的1 018.6 ind./haul升至1990s初期历史高值2 844.8 ind./haul,1990s后期剧降至42.07 ind./haul,2004—2015年AEDN徘徊在历史低值区间0.3~37.3 ind./haul (2004—2005年至历史低值0.3 ind./haul,不足1980s的0.03%);自2010s中后期始仔稚鱼AEDN呈明显恢复趋势,升至136.92~2 670.04 ind./haul区间(2017—2018年除外),至2020—2021年为历史次高值2 670.04 ind./haul(图 3d)。不同调查年份春夏季仔稚鱼AEDN差异极显著(Kruskal- Wallis H-Test for one-way ANOVA by ranks,H(10, n=213)= 72.32,P<0.001;各年均值比较χ2=64.39,df=10,P<0.001),呈先上升后下降而后又波动上升的趋势,由1982—1983年1 116.11 ind./haul升至1990s初期2 844.8 ind./haul,至1990s后期迅速降至42.07 ind./haul,2000s—2010s初期在历史低值区间0.62~5.64 ind./haul (2014—2015年至历史低值0.62 ind./haul);自2010s中期始,AEDN呈现波动上行趋势,至2020—2021年为历史高值5 424.75 ind./haul(图 3d)。不同调查年份夏季仔稚鱼AEDN差异显著(Kruskal-Wallis H-Test for one-way ANOVA by ranks,H(10, n=213)= 39.38,P<0.001;各年均值比较χ2=25.63,df=10,P<0.01),也呈先上升后下降而后又波动上升的趋势,由1982—1983年10.35 ind./haul升至2012—2013年75.21 ind./haul,此后在2013—2017年徘徊在2.31~16.56 ind./haul低水平区间,并在2014—2015年跌至历史低值2.31 ind./haul;自2010s后期开始,AEDN呈波动上行趋势,至2020—2021年达39.32 ind./haul (图 3d)。不同调查年份秋季AEDN均处0~2.19 ind./haul低水平区间,年际差异显著(Kruskal-Wallis H-Test for one-way ANOVA by ranks,H(9, n=175)=49.53,P<0.001;各年均值比较χ2=47.38,df=9,P<0.001),历史低值在2013—2014年,为0 ind./haul,当前2020—2021年为1.76 ind./haul (图 3d)。冬季AEDN年际差异显著(Kruskal-Wallis H-Test for one-way ANOVA by ranks,H(9, n=118)=48.71,P<0.001;各年均值比较χ2=35.89,df=9,P<0.001),历史低值在2017—2018年为0.4 ind./haul,历史高值为2014—2015年的106.9 ind./haul,当前2020—2021年为6.96 ind./haul (图 3d)。
2.3 渤海湾鱼类产卵(育幼)场分布年代际变动1980s渤海湾鱼卵EDN-ELH平面分布呈明显自湾口部至湾底部递增趋势,等EDN-ELH线与岸线呈近垂直状态,产卵场主要分布于118º00′E以西渤海湾中底部,核心产卵区(EDN-ELH高于5 000 ind./haul)位于潮白新河河口至漳卫新河河口间湾底部(图 4a)。1990s初期鳀大暴发,鳀卵高密度区(EDN-ELH高于1 000 ind./haul)几乎布满整个渤海湾,鱼卵EDN-ELH呈自湾东南口部至湾底部递增趋势(图 4b)。2000s—2010s初期,鱼卵EDN-ELH急剧下降,整个渤海湾无明显鱼类产卵场分布,特别是渤海湾底部自蓟运河河口至徒骇河河口间产卵场消失,鱼卵主要分布于湾东南口部近黄河入海口处(图 4c)。2010s中期开始,渤海湾鱼类产卵场呈恢复趋势,除自湾东北部突入的低密度区外,鱼卵EDN-ELH分布相对均匀,2个相对密度高值区分别位于潮白新河河口至漳卫新河河口间湾底部和东南部黄河入海口西北侧(图 4d)。2010s后期,渤海湾鱼卵EDN-ELH进一步升高,产卵场主要分布于118º00′E以西湾中底部、东南部近黄河入海口处和渤海湾东北口部,其中,湾底部潮白新河河口至漳卫新河河口间产卵场基本恢复(图 4e)。2020s初期,渤海湾鱼类产卵场规模进一步扩大,鱼卵EDN-ELH达历史峰值,高密度区布满整个渤海湾,整体呈现由湾口至底部递增的趋势(4f)。以118º30′E线为界,1982—1983、2016—2017和2020—2021年产卵重心在分界线以西的湾中底部;1998—1999、2004—2012、2014—2015和2018—2019年产卵重心在分界线以东的湾口部;1992—1993、2013—2014、2015—2016和2017—2018年重心分居分界线两侧(图 5a)。鱼卵分布重心由1980s位于渤海湾底部逐渐向湾口部迁移,至1990s末已迁移至118º30′E以东湾口部黄河入海口西北侧(图 5a);2000s—2010s初期,湾底部产卵场进一步衰退,产卵重心一直位于118º30′E以东湾口部(图 5a);2010s中期始产卵重心又迁回至118º30′E以西湾中底部(2018—2019年除外,位于118º30′E以东湾口部),其中,2016—2017年产卵重心位于118ºE以西湾底部(图 5a)。

|
图 4 1980s以来渤海湾各调查时期鱼卵生态密度分布 Fig.4 Variation of the distribution patterns to the spawning ground of fish in the Bohai Bay since the 1980s a: 1980s; b: 1990s; c: 2000s to pre-2010s; d: mid2010s; e: late 2010s; f: early 2020s |

|
图 5 1980s以来渤海湾各调查时期(年份)鱼卵(a)和仔稚鱼(b)分布重心迁移图 Fig.5 Migration trend of gravity center of fish eggs (a) and larvae (b) across different survey periods in the Bohai Bay since the 1980s |
1980s渤海湾仔稚鱼EDN-ELH高密度区(高于100 ind./haul)布满整个渤海湾,核心育幼场(高于500 ind./haul)位于独流减河河口至黄河故道刁口河口近岸海域及渤海湾口中部,并形成由渤海湾东北口部突入湾西南底部高密度区,而在渤海湾北部近岸海域密度相对较低(图 6a)。1990s渤海湾仔稚鱼EDN-ELH高密度区显著收缩,核心育幼场位于渤海湾北部海河口至滦河口近岸海域,此外渤海湾中底部密度也较高,存在自渤海中部和黄河入海口西北侧突入渤海湾低密度区(图 6b)。2000s—2010s初期,育幼场衰退明显,仔稚鱼EDN-ELH剧降,主要分布于渤海湾中底部,呈自湾口部至底部递增趋势,核心育幼场不明显(图 6c)。2010s中期,渤海湾育幼场呈现恢复,主要分布于湾中底部,并具一自渤海中部突入渤海湾中部低密度区,在独流减河河口至徒骇河河口间仔稚鱼密度较高(图 6d)。2010s后期,渤海湾仔稚鱼EDN-ELH进一步上升,育幼场主要分布于118º00′E以西湾中底部,核心育幼场分布于独流减河河口至徒骇河河口间渤海湾底部;存在自渤海湾东北口部突入中部的低密度区(图 6e)。2020s初期调查,鱼类育幼场规模已略高于1980s水平,核心育幼场布满整个渤海湾中底部;存在自黄河入海口西北侧突入渤海湾中部的低密度区(6f)。以118º30′E线为界,仔稚鱼分布重心除2004—2012和2018—2019年位于分界线以东湾口部外;其余年份重心主要分布于118º00′~118º30′E之间的湾中部,2020—2021年重心位于湾底部。

|
图 6 1980s以来渤海湾各调查时期仔稚鱼生态密度分布 Fig.6 Variation of the distribution patterns to the nursing areas of fish in the Bohai Bay since the 1980s a: 1980 s; b: 1990s; c: 2000 s to pre-2010 s; d: mid2010s; e: late2010s; f: early 2020s |
季节变化上,增温期春季和春夏季鱼卵优势和重要种类均以暖温性CPN鱼类为主,春夏季暖水性CPN鱼类、暖温或暖水性CD或CBD鱼类种数逐渐增多,部分年份春季暖水性CRA鱼类优势度也较高;夏季鱼卵优势或重要种类为暖水性或暖温性CD和CBD鱼类为主,部分年份暖温性CPN鱼类鳀为重要种;降温期秋季主要为CRA鱼类,冬季则未采集过浮性鱼卵(附录Ⅰ)。长期变化上,春季鱼卵优势和重要种类,1980s为暖温性小型CPN鱼类青鳞小沙丁鱼、斑鰶和CBD鱼类鮻(Planiliza haematocheilus)及暖水性CRA鱼类鲬(Platycephalus indicus);1990s前期更替为鳀,1990s后期为鳀、斑鰶和暖水性CPN鱼类赤鼻棱鳀;2000s—2010s初期年间变动比较剧烈,2004年为暖水性CBD鱼类小带鱼(Eupleurogrammus muticus)和暖温性CPN鱼类蓝点马鲛,2013—2014年更替为斑鰶和鲬,2014—2015年更替为鳀;2010s中期更替为鳀、鲬、青鳞小沙丁鱼和斑鰶;2010s后期更替为蓝点马鲛、鲬、斑鰶和鳀;2020s初又更替为鳀。长期变化上,春夏季优势和重要种类,1980s为暖温性CPN鱼类油魣(Sphyraena pinguis)、小带鱼、斑鰶和暖温性CD鱼类短吻红舌鳎(Cynoglossus joyneri);1990s初期更替为鳀、后期为鳀、斑鰶和赤鼻棱鳀;2000—2010s初期又更替为鳀、短吻红舌鳎、暖水性CBD鱼类叫姑鱼(Johnius grypotus)和赤鼻棱鳀;2010s中期更替不明显;2010s后期变为鳀、赤鼻棱鳀、短吻红舌鳎和暖水性CPN鱼类黄鲫;2020s初期更替为赤鼻棱鳀、短吻红舌鳎、小带鱼和青鳞小沙丁鱼(附录Ⅰ)。夏季优势和重要种类1980s为短吻红舌鳎、暖水性CBD鱼类白姑鱼(Pennahia argentata)和CD鱼类少鳞鱚(Sillago japonica);1990s初期为短吻红舌鳎,后期为小带鱼、短吻红舌鳎和少鳞鱚;2000—2010s初期为短吻红舌鳎、少鳞鱚、小带鱼和鳀;2010s中期以来,主要以短吻红舌鳎和少鳞鱚为优势或重要种类。长期变化,秋季优势和重要种类变化不大,各调查时期暖温性CRA鱼类花鲈处于绝对优势或重要地位,仅1998和2009年暖温性CD鱼类半滑舌鳎(Cynoglossus semilaevis)和鳀优势度上升成为优势或重要种类(附录Ⅰ)。
季节变化上,年内仔稚鱼在春季和春夏季优势和重要种类以暖温性CPN、CBD或CD鱼类为主,部分年份暖温性CRA鱼类花鲈、暖水性CPN鱼类赤鼻棱鳀和CD鱼类少鳞鱚优势度也较高;夏季暖水性CPN和CD鱼类优势度逐渐升高并在特定年份上升为优势种类,而暖温性CPN和CD鱼类仔稚鱼则是多数调查年份优势或重要种类,部分年份暖温性OEP鱼类尖嘴扁颌针鱼(Strongylura anastomella)优势度较高,为重要种类;秋季优势种年间变化较大,主要以暖温性CRA鱼类为主,部分时期暖温性或暖水性CPN鱼类及暖温性CD鱼类优势度较高;冬季以冷温性的CD鱼类为主,部分年份暖温性CRA鱼类花鲈或CPN鱼类沙氏下鱵(Hyporhamphus sajori)优势度较高,为优势或重要种类(附录Ⅱ)。长期变化上,春季仔稚鱼优势和重要种类自1980s至2010s初,暖温性小型CPN鱼类优势度呈下降趋势,暖温性小型CD和CBD鱼类优势度上升,至2020s初,暖温性小型CPN鱼类优势度略有恢复;春季仔稚鱼优势种类在1980s—1990s末期变化较大,由1980s的鳀变为1990s初期的花鲈和1990s末期的暖温性CD鱼类矛尾复鰕虎鱼(Acanthogobius hasta);自2000s—2010s末期均为暖温性CBD鱼类鮻,2020s初期又变为斑鰶(附录Ⅱ)。长期变化上,春夏季仔稚鱼优势和重要种类变动趋势类春季,1980s至2010s中期,小型CPN鱼类优势度降低,暖温性小型CD和CBD鱼类优势度上升,特别是1990s至2010s中期,隶属于暖温性CD鱼类的各类鰕虎鱼仔稚鱼如矛尾鰕虎鱼(Chaeturichthys stigmatias)、矛尾复鰕虎鱼和六丝钝尾鰕虎鱼(Amblychaeturichthys hexanema)等优势度明显上升,成为优势或重要种类;自2010s中期开始,渤海湾小型CPN鱼类优势度显著增高(附录Ⅱ)。长期变化上,夏季1980s—1990s末期无明显优势种类出现,1980s的重要种组成为尖嘴扁颌针鱼、少鳞鱚、赤鼻棱鳀、斑鰶和青鳞小沙丁鱼,1990s初期油魣阶段性迁入渤海湾成为重要种类,至1990s末期鰕虎鱼属鱼类优势度上升成为重要种类;自2010s初期开始,优势种显现,特别是产具卵膜丝卵的沙氏下鱵和暖水性CPN鱼类白氏银汉鱼(Hypoatherina valenciennei)等仔稚鱼优势度显著上升,成为各调查时期的优势或重要种类;自2010s中期以来,青鳞小沙丁鱼和鳀交替为渤海湾夏季仔稚鱼优势种类(附录Ⅱ)。长期变化上,渤海湾秋季仔稚鱼优势和重要种类年间和年代际更替明显,除1990s末期和2010s初外,花鲈均为各调查时期优势或重要种类;1982—1983年少鳞鱚、2014—2015和2018—2019年暖水性CPN鱼类如中颌棱鳀(Thryssa mystax)、赤鼻棱鳀和白氏银汉鱼等则为相应调查年份秋季重要种类(附录Ⅱ)。长期变化上,1980s—1990s末期,冬季仔稚鱼优势和重要种类年间变动显著,由1980s的花鲈变为1990s初期冷温性CD鱼类大泷六线鱼(Hexagrammos otakii)和1990s后期的冷温性CD鱼类玉筋鱼(Ammodytes personatus),自2013—2014年开始,除2018—2019年外,冷温性CD鱼类方氏云鳚(Pholis fangi)均为各调查年份优势种类(附录Ⅱ)。
2.5 渤海湾鱼卵、仔稚鱼物种多样性水平的季节变化和年代际变化多轮周年调查显示,渤海湾鱼卵H′ (因鱼卵DS变动趋势类H′,文中仅就H′展开分析)季节变化明显(Kruskal-Wallis单因素方差分析Kruskal-Wallis H-test for one-way ANOVA,H(4, n=55)=37.62,P<0.001;各季均值比较χ2=36.08,df=4,P<0.001)。表现为H′在增温期的春季和春夏季较高并呈上升趋势,年内最高值出现在春夏季;夏季H′开始下降,降温期秋季较低,冬季均未采集到过浮性鱼卵(图 7a)。春季鱼卵H′均值为0.86±0.49 (n=11),春夏季为1.06±0.48 (n=11)最高,夏季为0.83±0.24 (n=11)再次,秋季最低0.18±0.25 (n=11)。因鱼卵H′(图 8a)与DS (图 8b)变动趋势基本一致,本文将仅讨论H′变动,DS变动趋势这里不再赘述。长期变化上,春季鱼卵H′年间剧烈波动,在0.01~1.49区间,2014—2015年最低,2017—2018年最高(图 8a);H′高值点在1998—1999、2013—2014和2017—2018年;低值点在1992—1993、2014—2015和2018—2019年。1998—2015年H′逐年下降,由1998—1999年的1.18降至2014—2015年的0.01;2015—2018年又逐步升至历史高值1.49。春夏季鱼卵H′年间呈明显周期波动,在0.32~1.78区间,2012—2013年最低,1982—1983年最高(图 8a)。H′高值点在1982—1983、1998—1999、2013—2014、2014—2015、2016—2017、2017—2018和2020—2021年;低值点在1992—1993、2012—2013、2015—2016和2018—2019年(图 8a)。夏季鱼卵H′年间呈先波动上升,此后又波动下降的变化趋势,在0.38~1.31区间,2020—2021年最低,2014—2015年最高(图 8a)。H′低值点在1982—1983、2013—2014、2017—2018和2020—2021年。H′高值点在2014—2015和2016—2017年。1982—2015年H′波动上升,由1982—1983年0.73升至2014—2015年的1.31;此后H′波动下降至2020—2021年仅为0.38。秋季鱼卵H′均较低,呈先上升后下降而后又上升的变动趋势,在0~0.64区间(图 8a)。2009—2010年最高,2013—2014和2015—2019年各年份鱼卵H′均为0。1982—2010年H′处上升期,由1982—1983年的0.01升至2009—2010年历史高值0.64;在2010s,除2014—2015年H′为0.25外,其余各年H′均为0,2020—2021年为0.41(图 8a)。

|
图 7 渤海湾鱼卵(a)和仔稚鱼(b)香农–威纳指数(H′)季节变化特征箱型图
Fig.7 Box plot of the fish eggs (a) and larvae (b) Shannon-Wiener Index (H′) in different seasons
 平均值, 平均值, 平均值±标准误, 平均值±标准误, 平均值±标准差;数据源自表 1中所列自1980s来11个周期鱼卵仔稚鱼调查,n=11。
Mean, Mean±SE, Mean±SD; Data based on 11 round ichthyoplankton surveys carried out in the Bohai Bay since the 1980s lised in Tab.1, n=11. 平均值±标准差;数据源自表 1中所列自1980s来11个周期鱼卵仔稚鱼调查,n=11。
Mean, Mean±SE, Mean±SD; Data based on 11 round ichthyoplankton surveys carried out in the Bohai Bay since the 1980s lised in Tab.1, n=11.
|

|
图 8 渤海湾各调查时期不同季节鱼卵和仔稚鱼香农–威纳和辛普森多样性指数 Fig.8 Shannon-Wiener and Simpson's diversity index of fish eggs and larvae profiles along the survey season across different survey times in the Bohai Bay a和c分别为鱼卵和仔稚鱼香农–威纳指数;b和d分别为鱼卵和仔稚鱼辛普森多样性指数 Shannon-Wiener index of eggs (a) and larvae (c); Simpson's diversity index of eggs (b) and larvae (d) |
渤海湾仔稚鱼H′(因仔稚鱼DS变动趋势类H′,文中仅就仔稚鱼H′展开分析)季节变化显著(Kruskal-Wallis H-test for one-way ANOVA,H(4, n=55)= 18.6,P<0.001;各季均值比较χ2=16.44,df=4,P<0.01)。表现为H′在升温期的春季和春夏季逐次升高,至夏季升至年内峰值;在降温期秋季下降,冬季进一步降至年内谷值(图 7b)。春季仔稚鱼H′为0.58±0.33 (n=11),春夏季升至1.09±0.45 (n=11),夏季达年内高值1.22±0.49 (n=11),秋季降至0.64±0.59 (n=11),至冬季仔稚鱼H′最低0.39±0.35 (n=11) (图 7b)。因仔稚鱼H′ (图 8c)与DS(图 8d)变动趋势基本一致,本文将仅讨论仔稚鱼H′变动。长期变化,春季渤海湾仔稚鱼H′年间剧烈波动,在0.09~1.09区间,2004—2005年最高,2016—2017年最低。H′呈先上升后下降而后又上升的变动趋势(图 8c),高值点在1992—1993和2004—2005年;低值点在2014—2015和2016—2017年。1982—2005年H′波动上升,由1982—1983年的0.54上升至2004—2005年的1.09;2005—2017年H′波动下降至历史低值0.05;2010s后期始H′逐渐上升,至2020—2021年为0.63。春夏季H′年间剧烈波动,在0.39~1.89区间,2018—2019年最高,2020—2021年最低。H′呈先降后升、再降又升随后又下降变动趋势,1982—1999年H′呈下降趋势,1999—2016年波动上升,近年来波动加剧,2017—2018年急剧下降,随后2018—2019又升至历史高值,而2020—2021年又跌至历史谷值(图 8c)。夏季仔稚鱼H′呈先下降随后周期性波动趋势,在0.45~2.07区间(图 8c)。1992—1993年最高,2020—2021年最低。1982—1999年H′呈下降趋势,由1982—1983年1.78降至1998—1999年1.15;随后H′呈周期性波动,高值点在2013—2014、2016—2017和2018—2019年,低值点在2015—2016、2017—2018和2020—2021年。秋季H′波动剧烈,呈先下降后上升的变动趋势,为0~1.4 (图 7c),2016—2017年最高,1998—1999、2009—2010和2013—2014年均为零。1982—2014年H′呈下降趋势,由1982—1983年0.85和1992—1993年1.02降至0;随后自2014— 2015年开始,H′由0.27升至2016—2017年1.40历史高值,近年来一直维持在1.0以上。冬季H′年际间波动剧烈,在0~0.99区间,2013—2014年最高,1992— 1993年和2015—2016年均为0。自1982—2016年,H′由1982—1983年的0.17波动变为2015—2016年的0;此后,2016—2020年在0.23~0.60区间小幅振动(图 8c)。
2.6 近40年来渤海湾鱼类早期资源种类更替渤海湾各调查时期鱼类早期资源种类更替现象明显。1982—1983年与1992—1993年、2020—2021年鱼类早期资源物种组成CC值位于0.50~0.75区间,群落间物种组成中等相似;而与1998—1999、2004—2012、2013—2019年各年CC位于0.25~0.50区间,中等不相似(表 2)。1992—1993年与1982—1983年和1998—1999年CC值位于0.50~0.75区间,群落间物种组成中等相似;与其余年份CC值位于0.25~0.50区间,中等不相似(表 2)。1998—1999与1992—1993和2017—2018年CC值位于0.50~0.75区间,群落间物种组成中等相似;与其余调查年份CC值位于0.25~0.50区间,中等不相似(表 2)。2004—2012与2013—2014、2015—2019年CC值位于0.50~0.75区间,中等相似;而与其余调查年份CC值位于0.25~0.50区间,中等不相似。2013—2014年与2004—2012、2014—2018年各年CC值位于0.50~0.75区间,中等相似;而与其余调查年份CC值位于0.25~0.50区间,中等不相似(表 2)。2014—2015年与1982—1983、1992—1993、2004—2012、2017—2018和2020—2021年CC值位于0.25~0.50区间,中等不相似(表 2);而与其余调查年份CC值位于0.50~0.75区间,中等相似(表 2)。2015— 2016年与1982—1983、1992—1993和1998—1999年CC值位于0.25~0.50区间,中等不相似,而与其余年份CC值位于0.50~0.75区间,中等相似(表 2)。2016—2017年与1982—1999年间各调查年份CC值位于0.25~0.50区间,中等不相似;而与其余调查年份CC值位于0.50~0.75区间,中等相似(表 2)。2017—2018年与1982—1983、1992—1993和2014—2015年CC值位于0.25~0.50区间,中等不相似;而与其余年份CC值位于0.50~0.75区间,中等相似。2018—2019年与1982—1983、1992—1993和2013—2014年各年份CC值位于0.25~0.50区间,中等不相似,而与其余年份CC值位于0.50~0.75区间,中等相似(表 2)。群落间物种组成年间相似性程度由大到小排序为1998—1999、1982—1983、1992—1993、2020—2021、2015—2016、2018—2019、2016—2017、2017—2018、2004—2012、2013—2014、2014—2015年。
|
|
表 2 不同调查时期渤海湾鱼类早期资源物种组成年间β相似性指数矩阵 Tab.2 Matrix of the β similarity index (CC) of species composition to the ichthyoplankton community in the Bohai Bay between different survey periods |
1982—2021年研究的7种渤海湾鱼类鱼卵EDN-ELH存在明显的此消彼长现象。如果1982—1983年各鱼种鱼卵资源丰度指数为1,那么此后鳀卵EDN-ELH变动为0.92~65.37倍,黄鲫卵为1~3.24倍,蓝点马鲛卵为1~2.44倍,赤鼻棱鳀卵为1~2.24倍,斑鰶卵为0.44~1倍,青鳞小沙丁鱼卵为0.48~1.75倍,花鲈卵为0.41~1倍。变动幅度最大的为鳀,其次为黄鲫、蓝点马鲛、青鳞小沙丁鱼、赤鼻棱鳀、花鲈和斑鰶。40年来,鱼卵EDN-ELH明显降低的为斑鰶、青鳞小沙丁鱼和花鲈;明显升高的为蓝点马鲛、黄鲫、赤鼻棱鳀;鳀波动剧烈且呈升高趋势(图 9)。鳀EDN-ELH在1992—1993年达历史峰值65.37倍,此外,除2016—2017和2017—2018年AI低于1982—1983年外,其余年份均高于1982—1983年;其中,2015—2016年和2020—2021年为鳀高发年份,历史低值在2016—2017年(图 9)。黄鲫历史低值在1982—1983年,在1992—1993年达历史峰值3.24倍;1992—2021年间呈先下降后波动上行趋势,至2020—2021年达次高值1.81倍。蓝点马鲛AI历史低值在1982—1983年,历史峰值在2017—2018年为2.44倍;1992—1993年AI较1982—1983年显著升高,此后1998—2015年间蓝点马鲛AI处在相对较低区间,自2016—2021年明显上升(图 9)。赤鼻棱鳀AI历史低值在1982—1983年,历史峰值在2016—2017年,为1.81倍;自1982—1999年间呈上升趋势,1999—2014年呈下降趋势,2014—2017年呈明显上升趋势,而后又逐年下降,至2020—2021年为1.08倍(图 9)。斑鰶历史峰值在1982—1983年,1998—1999、2013—2014和2016—2017年为相对高值点;历史低值在2015—2016年仅0.48倍(图 9)。青鳞小沙丁鱼历史峰值在1992—1993年,为1.75倍,历史低值在2015—2016年,仅0.47倍;1982—1993年升至历史峰值,而后至1998—1999年急剧下降,自1999—2014年处在低值区间,2017—2020年间微弱上升(图 9)。花鲈历史峰值在1982—1983年,历史低值在2016—2017,仅0.41倍;自1992—2021年在低值区间内波动(图 9)。

|
图 9 1980s来渤海湾主要鱼类鱼卵资源丰度指数变动
Fig.9 Eggs abundance index of the main fish in the Bohai Bay since the 1980s
 斑鰶、 斑鰶、 青鳞小沙丁鱼、 青鳞小沙丁鱼、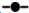 鳀、 鳀、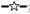 赤鼻棱鳀、 赤鼻棱鳀、 黄鲫、 黄鲫、 蓝点马鲛、 蓝点马鲛、 花鲈。
Konosirus punctatus, Sardinella zunasi, Thryssa kammalensis, Engraulis japonicas, Setipinna tenuifilis, Scomberomorus niphonius and Lateolabrax maculatus. 花鲈。
Konosirus punctatus, Sardinella zunasi, Thryssa kammalensis, Engraulis japonicas, Setipinna tenuifilis, Scomberomorus niphonius and Lateolabrax maculatus.
|
近40年来,不同调查时期渤海湾鱼类亲体各产卵类型种数均以产浮性卵种数最高,其次为产附着性卵、产黏着沉性卵和卵胎生鱼类,产凝集浮性卵鱼类仅在2020—2021年出现(图 10)。长期变化上,产浮性卵鱼类种数呈先下降后上升的变动趋势,种数由1982—1983年29种降至2014—2015年13种,2010s中期始种数逐渐增加,至2020—2021年升至20种;产附着性卵鱼类种数增加趋势比较明显,自1980s至2010s初期保持稳定在4~6种,自2010s中期始种数逐渐上升保持在9~12种,至2020—2021年为10种;产黏着沉性卵鱼类呈现微弱先降后升变动趋势,由1982—1983年3种降至2004—2012年的0种,此后波动上升,至2020—2021年为4种;卵胎生鱼类种数一直维持在1~3种(图 10)。长期变化,产浮性卵鱼类种数百分占比在各调查时期呈微弱先上升后波动下行的趋势,由1982—1983年的74.36%变为2004—2012年的77.27%,此后,降至55%~61.9%,2020—2021年为历史低值55%;产附着性卵鱼类百分占比呈明显上升趋势,由1982—1983年的12.82%升至2015—2016年的历史高值34.62%,2020—2021年为25%,百分占比升高1倍;产黏着沉性卵鱼类百分占比呈先下降后上升的变动趋势,由1982—1983年7.69%降至2004—2012年0,此后升至2020—2021年10% (图 10)。

|
图 10 1980s以来渤海湾不同调查时期亲体产卵类型年间种数比较 Fig.10 Comparison of the number of taxas in different spawning adaptation type of the spawning stock in the Bohai Bay between different survey periods since the 1980s P:产浮性卵鱼类;AF:产附着性卵鱼类;D:产黏着沉性卵鱼类;O:卵胎生鱼类;AP:产凝集浮性卵鱼类。 P: Pelagic egg; AF: Egg with adhesive filaments; D: Demersal egg; O: Ovoviviparity; AP: Agglutinative pelagic egg. |
40余年来,各调查时期渤海湾鱼类亲体栖息类型种数均以CD种数最高,其次为CPN、CBD、CRA鱼类,OEP种数年间波动较大,OMP鱼类仅在2020—2021年出现(图 11)。长期变化上,CD和CBD鱼类种数及其种数百分占比均呈先下降后上升的变动趋势。1982—2015年CD和CBD鱼类种数由1982—1983年27种降至2014—2015年11种,种数百分占比也由1982—1983年最高69.23%降至2014—2015年的52.38%;此后,CD和CBD种数逐步上升,至2020—2021年为26种,种数百分占比升至65%(图 11)。长期变化上,CPN鱼类种数保持相对稳定,除1990s为11种外,其余调查年份均在7~10种之间。CPN鱼类种数百分占比则呈先上升后下降的变动趋势,1982—1993年CPN鱼类种数百分比由1982—1983年的20.51%升至1992—1993年峰值36.67%;1993—2015年CPN鱼类种数百分占比均在30%以上(2013—2014年除外,为29.17%);从2010s中后期开始,CPN鱼类种数百分比降至25%~27.27%区间,2020—2021年为25% (图 11)。长期变化上,CRA鱼类种数2种左右,基本保持稳定(1992—1993和2015—2016年除外,仅1种);OEP鱼类[尖嘴扁颌针鱼和鲯鳅(Coryphaena hippurus)]年间波动较大;OMP种类[黄鮟鱇(Lophius litulon)]仅出现在2020—2021年5月调查中(图 11)。

|
图 11 1980s以来渤海湾不同调查时期亲体各栖息类型年间种数比较 Fig.11 Comparison of the number of taxas in different habitat type of the spawning stock in the Bohai Bay between different survey periods since the 1980s CD:大陆架浅水底层鱼类;CBD:大陆架浅水中底层鱼类;CPN:大陆架浅水中上层鱼类;CRA:大陆架岩礁性鱼类;OEP:大陆架大洋洄游性中上层鱼类;OMP:大洋深水底层鱼类。 CD: Continental shelf demersal fish; CBD: Continental shelf benthopelagic fish; CPN: Continental shelf pelagic-neritic fish; CRA: Continental shelf reef-associated fish; OEP: Oceanic pelagic fish; OMP: Oceanic bathydemersal fish. |
不同调查时期渤海湾鱼类亲体适温类型以WT鱼类最高,WW鱼类种数次之,CT鱼类种数最低,没有出现CW鱼类(图 12)。长期变化,WT和WW鱼类种数均呈先下降后上升的变动趋势,种数分别由1982—1983年26种和10种降至2014—2015年的12种和7种,此后种数逐渐上升,至2020—2021年分别为27种和11种;CT种数则一直在1~3种的较低区间。长期变化上,不同适温类型百分占比在各调查时期保持相对稳定,WT鱼类百分占比在各调查时期均超过55%,2018—2019年最高为69.70%,2014—2015年最低为57.14%,除1992—1993、1998—1999和2014—2015年外,WT值均在65%以上;WW鱼类百分占比为25%~33%,2013—2014年最低为25%,2014—2015年最高为33.33%,WW鱼类百分占比高值在1992—1993、1998—1999、2004—2012和2017—2018年,低值在1982—1983、2013—2014和2020—2021年(图 12);CT鱼类百分占比为2.9%~9.5%,1998—1999年最低为2.9%,2014—2015最高为9.5%,高值在1982—1983、2013—2014、2014—2015和2016—2017年,低值在1998—1999和2017—2018年(图 12)。

|
图 12 1980s以来渤海湾不同调查时期亲体适温类型年间种数比较 Fig.12 Comparison of the number of taxas in different temperature adaptation type of the spawning stock in the Bohai Bay between different survey periods since the 1980s CT:冷温种;WW:暖水种;WT:暖温种。 CT: Cold temperate species; WW: Warm water species; WT: Warm temperate species. |
渔业种群早期生活史阶段是其生命周期中最脆弱、对栖息环境变化敏感性最强的阶段,小规模环境变化也可能会对资源补充过程产生剧烈影响(Houde, 1987)。鱼类早期资源的存活和数量是渔业资源补充和可持续利用的基础(Houde, 1987),同时也是海洋环境变化的敏感指标(Nielsen et al, 2021)。挪威著名鱼类学家Hjort (1914)在20世纪初期提出了渔业资源评估最早的三个学说之一“波动论”,认为环境条件波动是形成鱼类世代强弱的主要原因。自Hjort (1914)的开创性研究以来,因对渔业种群动态的有限认知限制了人们做出有效渔业管理的能力,鱼类补充机制研究一直为渔业资源学研究热点(Somarakis et al, 2019)。近年来,许多研究也表明,捕捞活动可加剧渔业种群数量波动(Hsieh et al, 2006; Anderson et al, 2008),同时,全球气候变暖背景下主导渔业种群数量波动的环境驱动因子也常会发生转变(Rijnsdorp et al, 2009,Pörtner et al, 2010),但要将捕捞与气候变化两方面影响剥离非常困难。渤海湾历史上为多种鱼类如小黄鱼(Larimichthys polyactis) (夏世福, 1960; 刘效舜, 1960; 邱道立等, 1965; 朱崇俭等, 1965)、花鲈(吴光宗等, 1983; 万瑞景等, 1988)、半滑舌鳎(杨东莱等, 1984; 姜言伟等, 1988)、斑鰶和青鳞小沙丁鱼(孔立波等, 1996),虾类如中国对虾(Penaeus orientalis) (邓景耀, 1960、1980; 邓景耀等, 1983)和中国毛虾(Acetes chinensis) (辽宁省海洋水产研究所和河北省海洋水产研究所, 1961; 冯志青等, 1982、1987; “渤海湾、莱州湾毛虾适宜捕捞期的研究”课题组, 1992),蟹类如三疣梭子蟹(Portunus trituberculatus) (邓景耀等, 1988)和口足类如口虾蛄(Oratosquilla oratoria) (邓景耀等, 1988; 谷德贤等, 2018)等产卵场和其幼体主要索饵场。1980s,渤海湾为整个渤海卵子密度最大、产卵持续时间最长的海域(姜言伟等, 1988)。近40年来在过度捕捞、栖息地衰退和碎片化等共同作用下,渤海湾固有产卵场和索饵育肥场功能退化,一些重要渔业种群产卵场、索饵育肥场等栖息地丧失或发生变迁。
3.1 渤海湾鱼类早期资源结构的时间序列变化特征渤海湾鱼类以产浮性卵鱼类种数出现频率最高,其次为产附着性卵、产黏着沉性卵和卵胎生鱼类,偶见产凝集浮性卵鱼类;亲体栖息类型均以大陆架浅水底层鱼类种数最高,其次为大陆架浅水中上层、大陆架浅水中底层和大陆架岩礁性鱼类,大陆架大洋洄游性中上层鱼类种数年间波动较大,偶见大洋深水底层鱼类;亲体适温类型以暖温性鱼类种数最高,暖水性鱼类次之,冷温性鱼类种数最低。各调查时期鱼类早期资源的种类组成、优势种类、物种多样性水平和资源丰度等呈明显季节更替。鱼类产卵季节集中在5—8月升温阶段,5、6月进入渤海湾以产浮性卵为主的性成熟鱼类相继产卵,形成全年产卵高峰;7、8月水温全年最高,多数种类产卵期已过,鱼卵种类和数量明显减少,且由于沿岸河流入海径流流量增加,渤海湾盐度下降明显,有些已经在近岸浅水区开始或即将产卵种类移向渤海湾口部和外部(姜言伟等, 1988);10月湾内水温迅速下降,大多数洄游性种类相继游出渤海湾,整个调查海区鱼卵种类和密度均较低分布;11月至翌年3月,只有少数地方性(移动距离较短一般不洄游出渤海) (唐启升等, 1990; 邓景耀, 1988)产黏着沉性卵种类。小型CPN鱼类如斑鰶、青鳞小沙丁鱼、鳀、赤鼻棱鳀、黄鲫、中颌棱鳀、沙氏下鱵和白氏银汉鱼等;小型地方性CD和CBD鱼类,如矛尾鰕虎鱼、六丝钝尾鰕虎鱼、大银鱼(Protosalanx hyalocranius)、短吻红舌鳎、方氏云鳚和少鳞鱚等;大型地方性CD和CRA鱼类如鮻、半滑舌鳎、花鲈和鲬等为渤海湾各调查时期鱼类早期资源的主体成分。自1990s起,春季蓝点马鲛卵优势度显著提升,并在某些年份成为优势种类。有别于相邻莱州湾水域(卞晓东等, 2022b),渤海湾鱼卵H′均值在春夏季为年内最高值期,春季次之,夏季再次,秋季最低,冬季调查海区无浮性卵子分布;仔稚鱼H′均值在升温期的春季和春夏季逐次升高,夏季升至年内峰值,在降温期秋季下降,冬季最低。而莱州湾水域鱼卵H′在春季最高,春夏季次之,夏季为再次,秋季最低;仔稚鱼H′春夏季达年内高值,夏季次之,春季再次,冬季次低、秋季为年内最低。各调查季节渤海湾鱼卵H′与莱州湾鱼卵H′无明显差异[春季莱州湾(下同) H′=0.95±0.63(n=11),Mann-Whitney U Test, U=52,P>0.05;春夏季H′=0.91±0.61 (n=11),U=46,P>0.05;夏季H′=0.83±0.35 (n=11),U=50,P>0.05;秋季H′=0.15±0.24 (n=11),U=57.5, P>0.05]。渤海湾仔稚鱼H′在春季明显低于莱州湾[莱州湾仔稚鱼H′=1.12± 0.38 (n=11),Mann-Whitney U Test, U=14,P<0.01],而其他季节差异不显著[春夏季H′=1.52±0.45 (n=11),U=33,P>0.05;夏季H′=1.31±0.49(n=11),U=56,P>0.05;秋季H′=0.64±0.33(n=11),U=55,P>0.05;冬季H′=0.77±0.37 (n=11),U=31.5 P>0.05]。2020—2021年调查结果又显示,渤海湾鱼卵H′除6月略高于烟威近岸海域外,其余春季和夏季均低于烟威近岸海域;而仔稚鱼H′在春、春夏和夏季均低于烟威近岸海域(张雨轩等, 2022b)。渤海湾各鱼种生殖时期对温盐等水文要素适应条件不同,鱼类早期资源种类、优势种类和物种多样性水平的季节变化主要是不同生活史类型(产卵、适温和栖息类型)鱼类适应栖息地温盐等水文要素变化的结果。
自1980s以来,渤海湾鱼类早期资源结构处持续更替过程中,资源丰度呈激烈的年际和年代际波动。渤海湾鱼卵和仔稚鱼种类数和资源丰度整体呈先下降后上升的变动趋势,均在2010s初期跌至历史低值,近年来,特别自2017—2018年以来明显恢复。鱼卵和仔稚鱼种数由1980s的39种,1990s前期的31种、后期的34种,2000—2010s初期的22种,逐次降至2010s前期的21种,2010s中后期逐步回升,至2020—2021年已达40种。鱼卵种数和AI均在2014—2015年降至历史低值,种数不足1980s的50%,AI仅为1980s的20%;自2010s中期始鱼卵种数和AI值呈现恢复,至2020—2021年种数约为1980s的80%左右,AI约为彼时的88%。鱼卵AI在不同季节呈现不同的变动趋势,其中,春、春夏和夏季均呈先下降后上升的变动趋势,而秋季则呈显著下降趋势;相较1980s,当前鱼卵AI在春季和春夏季与其相当,夏季呈明显上升趋势,秋季下降明显。仔稚鱼种数和AI则均在2000—2010s初期降至历史低值,种数仅为1980s的37%,AI仅25%左右;自2010s中期开始,种数和AI呈现明显恢复状态,至2020—2021年仔稚鱼种数与1980s大致相当,AI则为彼时的1.33倍。仔稚鱼AI在春、春夏和夏季均呈先上升后下降随后波动上升的趋势,而秋季2013—2014年调查期间仔稚鱼密度极低,冬季2014—2015、2016—2017和1982—1983年显著高于其他年份;相较1980s,春季和春夏季仔稚鱼AI显著升高,夏、秋、冬季与1980s差异不显著。相较1980s,至2020—2021年渤海湾出现鱼类早期资源种类数和AI已与其无明显差异,但鱼类早期资源结构已发生明显变化。40余年来鱼类早期资源年间种类更替率均在26%~64%区间,即便是相邻年份间种类更替率也高达30%,且近年来呈明显加快趋势。主要产卵期内同季节鱼类早期资源优势种类更替现象明显,鱼卵和仔稚鱼H′呈年际和年代际剧烈波动。各产卵类型(产附着性卵鱼类除外)、栖息类型、适温类型产卵亲体种数大致呈先下降后上升的变动趋势,全年综合亲体产卵类型中产浮性卵种数显著下降,产附着性卵种数则显著升高;栖息类型中CPN鱼类所占比例升高,CD和CBD鱼类所占比例降低;适温类型中CT种数自2010s中期以来呈上升趋势。生命周期短、性成熟早、处食物链低端的小型CPN鱼类如鳀、赤鼻棱鳀和黄鲫,小型CD鱼类如矛尾鰕虎鱼、六丝钝尾鰕虎鱼、黄鳍刺鰕虎鱼(Acanthogobius flavimanus)和大颌裸身鰕虎鱼(Gymnogobius macrognathus)以及大型的CPN鱼类如蓝点马鲛等AI超过1980s水平。但渤海湾传统经济鱼类如小型CPN鱼类青鳞小沙丁鱼、斑鰶,大型CRA鱼类如花鲈,大型CD鱼类如半滑舌鳎等AI较1980s下降明显;特别是小黄鱼和带鱼等传统底层经济鱼类早期资源虽有出现,但EDN-ELH极低,近乎绝迹,表明渤海湾作为传统经济鱼类的产卵、育幼场所的功能有所下降。
3.2 渤海湾鱼类早期资源密度的空间变化特征渤海湾虽是半封闭海区,但它与外海水有着连续而强烈的交换;以外海高盐水系为主的黄海暖流余脉顺着黄海槽经渤海海峡进入渤海后分为两支,其中一支由渤海中部左转流向渤海湾,并与以沿岸低盐水系为主沿岸流形成逆时针环流(邓景耀等, 1983)。各调查时期渤海湾鱼类产卵场和索饵场一般集中分布于湾中底部两种不同性质海流交汇区亦即两种不同水系混合区,由于在此区域营养盐较丰富,饵料生物繁生,是鱼类繁育后代的良好场所(中华人民共和国黄海水产研究所, 1960)。因渤海湾多数洄游鱼类从越冬场到产卵场进行生殖洄游期间对盐度的适应是一个从高盐区逐渐过渡到低盐区的过程,当由暖流和沿岸流年际强弱变化及其消长而引起不同水系交汇区位置和盐度发生剧变时,往往会影响亲体产卵行动从而引起产卵场位置变化(中华人民共和国黄海水产研究所, 1960)。又因自然海域鱼类早期生活史阶段(浮性卵和初孵仔鱼)主营浮游生活,特别是浮性卵完全没有自主游泳能力,在风、流、波浪等作用下随波逐流地漂移。渤海湾中底部海水的流动主要由随潮汐周期流动的潮流所构成,受地形所致,湾内潮流向基本属东西向(邓景耀等, 1983),所以各调查时期湾中底部鱼卵等EDN-ELH线排列为顺着东西向潮流近与岸线垂直,仔稚鱼等EDN-ELH线排列趋势亦然。
1980s,渤海湾产卵高峰期(5—6月)除鳀、鲬和蓝点马鲛等少数种类在渤海湾口部及外部盐度和透明度较高水域产卵外,多数种类如优势种斑鰶和青鳞小沙丁鱼等先后进入渤海湾底部受沿岸低盐水系控制浅水河口和附近海区产卵繁殖,产卵重心位于海河口以南至捷地减河河口以北水深5~13 m海区(姜言伟等, 1988)。1990s,随着黄河(吴德星等, 2004; 东营市水利志编纂委员会, 2003)和海河(雷坤等, 2007)等入海径流激减和黄河经常性断流,各河口附近入海径流冲淡过程减弱,低盐区缩小,受盐度梯度驱动物理潮汐和湾口环流退化并开始转向,外海高盐水入侵加强,渤海湾及湾口盐度普遍升高(吴德星等, 2004; 雷坤等, 2007; Wang et al, 2010; Wu et al, 2020)。如1961—1996年的35年中,湾底部受沿岸低盐水系控制塘沽站年平均盐度增加1.9,湾东南部老黄河口外海表层的低盐区已由高盐区替代(吴德星等, 2004)。相应1990s初期,适应盐度和透明度较大的鳀(卞晓东等, 2022a)卵子扩大其分布范围至整个渤海湾,数量在调查中占绝对优势,而斑鰶和青鳞小沙丁鱼卵子和仔稚鱼数量激减,仅分别为1980s的12.4%和65.5% (万瑞景等, 1998),渤海湾产卵重心也外迁至湾中部盐度较高的118º30′E线附近;1990s后期,随着黄河和海河等入海径流量进一步下降,渤海湾密集产卵区已不明显,产卵重心迁移至湾口东南部老黄河口外海高盐区域,仍以鳀卵为优势种。进入21世纪,整个2000s,对鱼类产卵繁殖有重要意义的春季(3—5月)黄河年均入海径流量降至历史低值14.87亿m3 (水利部黄河水利委员会, 2002—2021),湾底部产卵场进一步衰退,鱼卵AI也跌至历史低点(未考虑捕捞因素),产卵适盐和透明度较高的蓝点马鲛优势度上升,产卵重心仍位于湾口部。2010s初期,自2012年起,春季黄河入海径流量回升,渤海湾鱼卵AI仍维持在低位,产卵重心一直在118º30′E线以东湾口部,其中,PDO指数冷位相和春季黄河入海径流量相对充沛的2013—2014年(卞晓东等, 2022a)优势种为斑鰶,重心靠近118º30′E线;而PDO指数暖位相和春季黄河入海径流量相对较少的2014—2015年(水利部黄河水利委员会, 2002—2021)优势种为鳀,重心偏向湾口部。2010s中期,渤海湾产卵重心也呈激烈年际变化,产卵适温较斑鰶高(产卵期晚)的近岸内湾产卵赤鼻棱鳀(卞晓东等, 2022a)扩大其分布范围且优势度上升;尽管2016—2017年春季黄河入海径流量相对较少(水利部黄河水利委员会, 2002—2021年),但当年鳀发生量较少,优势种为斑鰶,产卵重心位于118ºE以西湾底部;2017—2018年5月,产卵适盐和透明度较高的蓝点马鲛和鲬为优势种类,6月鳀(主要分布于渤海湾119º00′E以东湾口部)和赤鼻棱鳀(主要分布于渤海湾118º35′E以西湾中部)为优势种类,产卵重心位于118º30′E线附近;2018—2019年春季黄河入海径流量为1990s初期以来的最高值(水利部黄河水利委员会, 2002—2021),但因调查站位主要布设于118º00′E以东盐度和透明度较大区域(湾底部无站位),当年5、6月调查优势种仍然为鳀。进入2020s,春季黄河入海径流量达48.55亿m3的历史高位(水利部黄河水利委员会, 2002—2021),5月鳀为优势种广泛分布于118º00′E线以东湾中部至湾口部海域,赤鼻棱鳀为重要种类分布于118º15′E线以西湾中底部;6月赤鼻棱鳀为优势种主要分布于118º00′E以东海域,全年产卵重心位于118ºE线附近。
3.3 渤海湾鱼类早期资源动态变化的“上行控制”因素初析渤海湾为一个三面环陆的浅水海湾,气象条件对水文要素的变化影响巨大(农业部水产局和农业部黄海区渔业指挥部, 1990)、生态环境脆弱,是气候变化影响的敏感区域(自然资源部国家海洋信息中心, 2020—2022)。具有激烈年变化的渤海湾水温特点及其各种水文要素在空间分布和时间变化上的特征及规律,在相当大程度上决定了渤海湾产卵场范围大小、进入产卵场的种类和数量以及鱼类早期资源密度分布。水温和盐度是影响海洋硬骨鱼类早期资源发育和存活的2个最主要的环境因子(Bunn et al, 2002),水温控制鱼类生理生化进程,盐度控制鱼类渗透调节,进而使鱼类的生理机能发生变化,但在自然条件下很难把温度和盐度因素截然分开,从而找出影响生物个体各发育阶段的主导因素。全球气候变暖背景下,自1960s以来,渤海湾年均海表温(SST)呈显著波动上升趋势,其中1982—1997年处在升温期(warming period);1998—2013年尽管处于升温间歇期(warming hiatus period),但渤海湾全年暖昼日数和极端高温事件累积强度均呈现上升趋势(Li et al, 2019、2021);2015—2020年渤海湾年均SST连续6年处于高位;自1980s以来,渤海湾海洋热浪发生频次、持续时间和累积强度均呈显著增加趋势,特别自2019年以来,渤海湾发生海洋热浪频次年均在7次以上,海洋热浪时间年均80 d以上,平均热浪强度在1.2 ℃以上(自然资源部国家海洋信自然资源部国家海洋信息中心, 2020—2022)。海洋升温会改变产卵育幼场的环境条件,将影响受精卵的孵化率、发育速率和孵化期长短,初孵仔鱼的发育状态、营养物质利用效率及其散布潜能(Pepin et al, 1997; Laurel et al, 2008; Pinsky et al, 2013; Bian et al, 2014、2016; Free et al, 2019),改变饵料生物可获得性(Beaugrand et al, 2018),直接决定鱼类早期资源存活状况,进而影响世代强弱(Almatar, 1984)。海洋热浪频发则会导致海洋生态系统鱼类早期资源结构的变化。在2014—2016年间频发的海洋热浪已导致东北太平洋沿岸鱼类早期资源结构发生深刻变化,如在高纬度阿拉斯加湾和加拿大不列颠哥伦比亚省沿岸海域,多数传统优势类群早期资源AI处于自采样记录来的最低水平;而在与渤海湾纬度相近美国俄勒冈州沿岸和南加州洋流区,暖水性鱼类早期资源物种丰富度和AI呈上升趋势;且与厄尔尼诺事件影响明显不同,海洋热浪频发年份在南加州沿岸海域冷温性鱼类早期资源AI也呈上升趋势(Nielsen et al, 2021)。在年均SST处高位、海洋热浪频发的2015—2020年,渤海湾海域少鳞鱚和白氏银汉鱼等暖水性鱼类EDN-ELH显著升高,冬季冷温性鱼类方氏云鳚等早期资源AI也呈上升趋势(附录Ⅱ)。
盐度的高低变化将直接影响卵子浮性和仔稚鱼渗透调节(Holliday et al, 1960; Laurence et al, 1981)。温盐组合的影响特别是水温被认为是限制产卵场地理分布的主要因子,而温度影响又被盐度效应所调控,特别是在接近产卵场地理分布边界(适温阈值)时,盐度效应尤为显著,表现为低温低盐和高温高盐增效作用(较低产卵场水温叠加较低盐度,或较高水温叠加较高盐度,有助于提高潜在补充成功率);相反,低温高盐和高温低盐抑制作用(较低水温叠加较高盐度,或较高水温叠加较低盐度,将降低潜在补充成功率) (Alderdice et al, 1971; Bunn et al, 2002; Bian et al, 2016; 卞晓东等, 2022a)。渤海湾产卵场盐度变化与沿岸降水量和河流入海径流量大小有密切关系(邓景耀等, 1983)。因各河流入海径流量具明显的季节变化,渤海湾产卵场盐度由沿岸季节性低盐水和外海高盐水相互影响,交替控制。1960s以来,海河水系降水量减少(邹磊等, 2021)、天然径流偏低等自然因素,叠加流域人类活动如水库和防潮闸建设及沿河取水取沙等,导致海河入海径流量已基本上是汛期径流(每年10月至翌年5月非汛期防潮闸关闭多数河流断流) (雷坤等, 2007),汛期外渤海湾产卵场盐度变化主要由黄河入海径流量变动引起(吴德星等, 2004; 雷坤等, 2007)。对渤海湾洄游性鱼类产卵繁殖具有重要意义的春季黄河入海径流量的年际变异会影响渤海湾产卵场范围大小、进入产卵场的种类和数量,进而影响鱼类早期资源密度。自1970s至2000s初期,黄河春季入海径流量一直呈负增长,由1970s的40.39亿m3激降至1980s的23.17亿m3、1990s的15.54亿m3和2000s的14.87亿m3,自2010s中后期起回升至31.08亿m3,至2020年为48.55亿m3(东营市水利志编纂委员会, 2002; 水利部黄河水利委员会, 2002—2021),呈明显的先下降后上升的趋势。这与渤海湾鱼类早期资源种类数和AI指数变动基本一致。随着河口海区冲淡过程减弱,盐度上升,河口海区形成渤海湾主要经济生物如小黄鱼(夏世福, 1960; 邱道立等, 1965; 朱崇俭等, 1965)、斑鰶和青鳞小沙丁鱼(孔立波等, 1996; 卞晓东等, 2022a)和中国对虾(邓景耀, 1960)等产卵场条件之一的低盐特性已发生改变,产卵场面积将随之缩小,从而影响到上述种类后代幼体的存活和补充;与此同时,生殖期适应盐度和透明度较高种类如鳀(卞晓东等, 2022a)、蓝点马鲛(卞晓东等, 2022a)和带鱼(夏世福, 1960)等将因生殖期河水流量的减少而可能扩大其分布范围,或移往河口海区的高盐区域,将有利于此类鱼类资源增殖。如自1990s以来,随着渤海湾盐度不断上升(吴德星等, 2004; 雷坤等, 2007),蓝点马鲛产卵场范围已扩展至118º30′E以西湾底部(农业农村部, 2021),在黄河入海径流量明显偏低,1992—1993年鳀卵更是布满整个渤海湾,1992年8月在渤海湾底部则采到了带鱼卵等(万瑞景等, 1998);同期斑鰶和青鳞小沙丁鱼等产卵场范围和EDN-ELH显著降低,特别是小黄鱼卵子在2010s以来的调查中近乎绝迹,当下渤海湾内已无明显小黄鱼产卵场分布。自2002年来,黄河调水调沙工程开始实施,使得黄河淡水和营养盐在年内输送更加集中,黄河入海物质通量季节性模式发生变化(Wang et al, 2010; 巩瑶, 2012; 龙跃等, 2017; Wu et al, 2020)。这个工程的实施为河口海域带来充足淡水和营养物质,并维持产卵场低盐水体特性的同时,又限制了繁殖适盐较高、产卵期与现行调水调沙期基本一致鱼类如带鱼等在黄河冲淡水影响区域(莱州湾黄河口海域和渤海湾等)的产卵繁殖活动。如自2000s以来,渤海湾海域调查中也鲜有带鱼卵子采集,当下渤海湾内已无明显带鱼产卵场分布。
3.4 捕捞对渤海湾鱼类早期资源动态变化的影响生殖群体和鱼类早期资源是渤海湾渔业生产和资源可持续利用的基础,捕捞强度和捕捞方式均会对其产生重要影响(邓景耀等, 1988)。如1950s,渤海湾渔业生产以群众渔业的近岸小风船为主,生产网具为扒拉网、风网、流网和转轴网等为主(天津市水产局区划办公室, 1980),主要捕捞利用资源生殖群体。渤海湾小黄鱼、带鱼等经济鱼类在集群与洄游分布上具明显季节特点,通常于生殖期集中于湾内产卵,集群密度高,易于捕捞,并形成群众渔业主要捕捞对象(刘效舜, 1960);当时小黄鱼卵和仔稚鱼广泛分布于渤海湾118º30′E以西湾底部(夏世福, 1960)以及东北部大清河口附近(朱崇俭等, 1965),卵最高密度达1 000粒/网,在多种鱼类卵子数量上占绝对优势。自1960s初至1970s末,尽管渤海捕捞强度(fishing effort in the Bohai Sea, FEBH)维持在较低水平(以年总渔获量annual total catch ATC和年海洋渔业机动渔船主机总功率数annual total break horsepower of the motor fishing vessels ATHPF来表征) (卞晓东等, 2022a),但随着机动渔船增加,底拖网渔业兴起,小黄鱼、带鱼等成鱼和幼鱼的越冬群体又遭受滥捕;自1960s初期在渤海开始的秋汛捕对虾,又兼捕了大量小黄鱼和带鱼等经济鱼类幼鱼(《中国渔业资源调查和区划》编辑委员会, 1988)。至1970s中后期,渤海小黄鱼和带鱼等传统底鱼资源已处于衰退或严重衰退阶段,其鱼群已形成不了渔业捕捞标志,如洄游路线、渔场和渔期等已不明显,逐渐失去其重要渔业地位(《中国渔业资源调查和区划》编辑委员会, 1990);至1980s初期,渤海湾小黄鱼产卵场分布范围已显著缩小,仅在118º00′E以西湾底部海河口至岐河口附近海域出现少量鱼卵,同期的一些低质小型中上层鱼类如青鳞小沙丁鱼、斑鰶、赤鼻棱鳀和鳀等EDN-ELH相对上升并成为优势种类(姜言伟等, 1988)。1980s中期至1990s末,为FEBH增长最为迅速阶段(1985—1998年ATHPF保持高速持续增长,ATC在1986—1992年增长,1992年急剧下降,而后逐步上升并在1998年达峰值) (卞晓东等, 2022a),并伴随着新增机动渔船小型化发展趋势;渤海湾小型机动渔船数量激增,其主要在渤海湾内产卵、育幼场密集的沿岸或河口海区生产,湾内小型流动渔具和定置渔具也随之激增(《中国渔业资源调查和区划》编辑委员会, 1990)。捕捞力量恶性膨胀直接导致渤海湾内传统渔业种类如小黄鱼、带鱼、花鲈、半滑舌鳎、青鳞小沙丁鱼和斑鰶等生殖群体资源量进一步下降(补充型捕捞过度);同时渤海湾沿岸定置网渔业在渔业生产中也占据重要地位(邓景耀, 1988; 孙中之等, 2012),定置网渔业生产过程中又兼捕了大量的经济鱼、虾类早期资源(生长型捕捞过度) (辽宁省海洋水产研究所和河北省海洋水产研究所, 1961; 冯志青等, 1982、1987; “渤海湾、莱州湾毛虾适宜捕捞期的研究”课题组, 1992),使渤海湾渔业资源遭到了生长型和补充型的双重捕捞过度,破坏了渔业资源可持续能力(农业部水产局和农业部黄渤海渔业指挥部, 1990);至1990s后期,渤海湾传统捕捞种类如小黄鱼、花鲈、半滑舌鳎、青鳞小沙丁鱼和斑鰶等鱼类早期资源AI均急剧下降,特别是小黄鱼和带鱼等经济鱼类产卵场已经不显著(程济生, 2004; 卞晓东等, 2018)。2000—2010s中期,渤海ATHPF依旧保持在高位,但ATC自2000—2008年间出现下降趋势,2008—2015年则进入平台期;自2015年起,FEBH (ATC和ATHPF)则均呈下降趋势(卞晓东等, 2022a)。期间,渤海湾鱼类早期资源种类数和资源丰度持续降低,均在2014—2015年跌至历史最低水平;2010s中期始开始恢复,至2020s初,种类数和资源丰度已与1980s大致相当,但半滑舌鳎和花鲈等底层经济鱼类EDN-ELH仍处历史低位。
国家渔业行政主管部门自1988年起实施渤海全年禁止底拖网作业,并相继开展了限制渤海沿岸海域定置网具数量、作业种类及水深,发展流网和钓钩渔业等捕捞作业方式优化调整措施(《中国渔业资源调查和区划》编辑委员会, 1988);自1995年起又开始在渤海海域实施“海洋伏季休渔制度”,“休渔期”又经历了从短到长的过程(卞晓东等, 2022a)。上述举措均为影响FEBH的重要因素,特别是“休渔期”起迄时间及时长又会对产卵期和传统渔期各异的渤海湾鱼类EDN-ELH动态产生影响。如传统渔期时长较长、开始时间较早,但渤海湾内产卵期相对较晚(5月下旬至6月中旬)的蓝点马鲛EDN-ELH自1980s至2010s中期一直处于相对稳定的状态,其关键在于黄渤海蓝点马鲛捕捞普遍采用的流网作业方式,因流网网目(较大)受到一定限制,这就使得相当数量的补充群体和剩余群体得到保护(韦晟, 1981);自1990s中后期开始施行的“伏季休渔制度”则通过直接关闭渔业,保护了大量蓝点马鲛产卵亲体和当年生幼体,从而使其后代能够延续和补充;随着“休渔期”起迄时间不断提前及时长延长,如自2017年开始,起始日期被提前到5月1日,时长延长至4个月,蓝点马鲛产卵期和其在渤海传统渔期均已涵盖在“休渔期”内,这样对产卵亲体捕捞压力大为减轻,2017—2018年蓝点马鲛EDN-ELH又较2016—2017年显著提升(图 9)。产卵期一直在“休渔期”外的鱼类,如花鲈(生殖期为8月上旬到11月上旬,10月上旬到10月下旬为生殖盛期) (万瑞景等, 1988)和半滑舌鳎(产卵初期始于8月下旬,结束于10月上旬,盛期为9月上中旬) (姜言伟等, 1988)等,自1980s中期以来一直遭受巨大捕捞压力,其EDN-ELH一直维持在较低水平。产卵期和传统渔期相对较晚,一直在“休渔期”内种类如少鳞鱚和赤鼻棱鳀等EDN-ELH自1990s以来一直处于相对稳定的状态。鮻大部分时间栖息于沿海浅水、礁石及海藻丛生海区,4月底至5月初在渤海湾各河口区产卵,除越冬期外,鮻一般不结大群;对鮻集中捕捞也仅在2—3月开凌期,其他时间多以定置网或小型网具生产,捕捞强度不大;近20余年来沿海海藻养殖业的发展,又为鮻创造了良好的隐蔽栖息场所(农业部水产局和农业部黄渤海渔业指挥部, 1990),因而其一直为各调查时期春季仔稚鱼的重要种类。
4 小结40余年来,渤海湾鱼类早期资源结构处于持续更替过程中,不同时期早期资源的种类组成、资源丰度、优势种类和物种多样性水平等呈明显季节更替;渤海湾产卵和育幼场范围和重心位置也处于持续更替过程中,渤海湾中底部均为各调查时期产卵场和育幼场集中分布区。相较1980s,渤海湾鱼类早期资源已发生结构性改变,渤海湾作为传统经济鱼类的产卵和育幼场所功能下降明显。传统经济鱼类如斑鰶、青鳞小沙丁鱼、小黄鱼、带鱼、花鲈和半滑舌鳎等EDN-ELH显著下降;繁殖期相对较早且产卵适应盐度和透明度较大的鱼类如鳀和蓝点马鲛,生殖期相对较晚的暖水性鱼类如赤鼻棱鳀和少鳞鱚等EDN-ELH则显著升高。全球变暖背景下渤海湾海表温持续上升和海洋热浪频发,以及黄河和海河等入海径流量减少而引起的渤海湾盐度升高和产卵场条件恶化;高捕捞强度减少了参加产卵的亲体量,网目尺寸不合规范的禁用渔具使用以及产卵期内捕捞利用等对鱼类早期资源的损害,均对渤海湾鱼类早期资源发生产生了重要影响。但要将捕捞与气候变化两方面对渤海湾鱼类早期资源的影响剥离开来非常困难。渤海湾鱼类早期资源结构及其动态变化是环境和捕捞双重扰动下的鱼类群落内生态位错位交替和结构性渔业资源衰退的具体体现。
附录Ⅰ 1980s来渤海湾各调查时期鱼卵优势(IRI>1 000)和重要(IRI>200)种类及其亲体适温和栖息类型
Appendix Ⅰ The dominant (IRI>1 000) and important (IRI>200) taxas of fish eggs with temperature adaptation (TA) and habitat types (HT) to the spawning stock in the Bohai Bay since the 1980s
附录Ⅱ 1980s来渤海湾各调查时期仔稚鱼优势(IRI>1 000)和重要(IRI>100)种类及其亲体适温、栖息和产卵类型
Appendix Ⅱ The dominant (IRI>1 000) and important (IRI>100) taxas of fish larvae with temperature adaptation (TA), habitat types (HT) and spawning types (ST) to the spawning stock in the Bohai Bay since the 1980s
http://journal.yykxjz.cn//uploadfile/yykxjz/20230914/Bian.pdf
致谢: 对各航次海上调查期间帮助采样的黄海水产研究所资源室各位前辈和科研人员谨致谢忱,感谢项目助理张晓燕同志在资料归集等方面提供的帮助,同时感谢国家农业科学渔业资源环境青岛观测实验站对本项研究给予的资助!
ALDERDICE D F, FORRESTER C R. Effects of salinity, temperature, and dissolved oxygen on early development of the Pacific cod (Gadus macrocephalus). Journal of the Fisheries Research Board of Canada, 1971, 28: 883-902 DOI:10.1139/f71-130 |
ALMATAR S M. Effects of acute changes in temperature and salinity on the oxygen uptake of larvae of herring (Clupea harengus) and plaice (Pleuronectes platessa). Marine Biology, 1984, 80: 1432-1793 |
ANDERSON C N K, HSIEH C, SANDIN S A, et al. Why fishing magnifies fluctuations in fish abundance. Nature, 2008, 452(7189): 835-839 DOI:10.1038/nature06851 |
BAI Y C, SHI F S, XU H J, et al. Coastline and tidal current changes responses due to large-scale reclamation in Bohai Sea. Marine Science Bulletin, 2021, 40(6): 621-635 [白玉川, 史丰硕, 徐海珏, 等. 渤海湾大规模围填海导致的岸线变化及潮流场响应分析. 海洋通报, 2021, 40(6): 621-635 DOI:10.11840/j.issn.1001-6392.2021.06.003] |
BEAUGRAND G, KIRBY R R. How do marine pelagic species respond to climate change? Theories and observations. Annual Review in Marine Science, 2018, 10: 169-197 DOI:10.1146/annurev-marine-121916-063304 |
BIAN X D, WAN R J, JIN X S, et al. Early life resources assemblage structure and succession to the marine Osteichthyes in the Laizhou Bay of Bohai Sea. Journal of Fisheries of China, 2022b, 46(1): 51-72 [卞晓东, 万瑞景, 金显仕, 等. 渤海莱州湾硬骨鱼类早期资源群落结构及演变. 水产学报, 2022b, 46(1): 51-72] |
BIAN X D, WAN R J, JIN X S, et al. Ichthyoplankton succession and assemblage structure in the Bohai Sea during the past 30 years since the 1980s. Progress in Fishery Sciences, 2018, 39(2): 1-15 [卞晓东, 万瑞景, 金显仕, 等. 近30年渤海鱼类种群早期补充群体群聚特性和结构更替. 渔业科学进展, 2018, 39(2): 1-15] |
BIAN X D, WAN R J, SHAN X J, et al. Preliminary analysis on recruitment variation and the exogenous driving factors to early life stages of the small pelagic fish in the Laizhou Bay. Journal of Fishery Sciences of China, 2022a, 29(3): 446-468 [卞晓东, 万瑞景, 单秀娟, 等. 莱州湾中上层小型鱼类早期资源量动态及其外在驱动因素. 中国水产科学, 2022a, 29(3): 446-468] |
BIAN X, ZHANG X, SAKRAI Y, et al. Temperature-mediated survival, development and hatching variation of Pacific cod Gadus microcephalus eggs. Journal of Fish Biology, 2014, 84: 85-101 DOI:10.1111/jfb.12257 |
BIAN X, ZHANG X, SAKRAIY, et al. Interactive effects of incubation temperature and salinity on the early life stages of Pacific cod Gadus microcephalus. Deep Sea Research Part Ⅱ, 2016, 124: 117-128 DOI:10.1016/j.dsr2.2015.01.019 |
BUNN N A, FOX C J, WEBB T. A literature review of studies on fish egg mortality: Implications for the estimation of spawning stock biomass by the annual egg production method. Science Series Technical Report, the Center for Environment, Fisheries and Aquaculture Science, Lowestoft, UK, 2000, 111: 1–37
|
CHENG J S, QIU S Y, LI P J, et al. Ecological environment and biotic community in the coastal waters of the Yellow Sea and Bohai Sea. Qingdao: China Ocean University Press, 2004: 343-350 [程济生, 邱盛尧, 李培军, 等. 黄渤海近岸水域生态环境与生物群落. 青岛: 中国海洋大学出版社, 2004: 343-350]
|
DENG J Y, KANG Y D, JIANG Y W, et al. A summary of surveys of the penaeid shrimp spawning ground in the Bohai Bay. Marine Fisheries Research, 1983, 5: 17-32 [邓景耀, 康元德, 姜言伟, 等. 渤海湾对虾产卵场调查. 海洋水产研究, 1983, 5: 17-32] |
DENG J Y, ZHU J S, CHEN J S, et al. Main invertebrates in the Bohai Sea and their fishery biology. Marine Fisheries Research, 1988, 9: 91-120 [邓景耀, 朱金声, 程济生, 等. 渤海主要无脊椎动物及其渔业生物学. 海洋水产研究, 1988, 9: 91-120] |
DENG J Y. Ecological bases of marine ranching and management in the Bohai Sea. Marine Fisheries Research, 1988, 9: 1-10 [邓景耀. 渤海渔业资源增殖与管理的生态学基础. 海洋水产学研究, 1988, 9: 1-10] |
DENG J Y. Preliminary investigation results of hatching conditions to Fenneropenaeus chinensis in sea area (Investigation Report No.58 of the Yellow Sea Fisheries Research Institute, Ministry of Fisheries P.R.China). Yellow Sea Fisheries Research Series, 1960, 2: 68-72 [邓景耀. 自然海区对虾孵化条件的初步调查结果(中华人民共和国水产部黄海水产研究所调查研究报告第58号). 黄海水产研究丛刊, 1960, 2: 68-72] |
DENG J Y. Distribution of eggs and larvae of penaeid shrimp, Penaeus orientalis, in Bohai Bay and its relation to natural environment. Marine Fisheries Research, 1980, 1: 17-25 [邓景耀. 渤海湾对虾(Penaeus orientalis)卵子、幼体数量分布及其与外界环境的关系. 海洋水产研究, 1980, 1: 17-25] |
Dongying Water Conservancy Records Compilation Committee. Dongying Water Conservancy Records (2002). Beijing: Hongqi Press, 2003: 56-57 [东营市水利志编纂委员会. 东营市水利志(2002). 北京: 红旗出版社, 2003: 56-57]
|
Editorial Committee of Fishery Resources Survey and Regionalization in China. Fishery resources survey and regionalization in China (Ⅰ): Regionalization of fishery. Hangzhou: Zhejiang Science and Technology Press, 1988: 155-158 [《中国渔业资源调查和区划》编辑委员会. 中国渔业资源调查和区划之一中国渔业区划. 杭州: 浙江科学技术出版社, 1988: 155-158]
|
Editorial Committee of Fishery Resources Survey and Regionalization in China. Fishery resources survey and regionalization in China (Ⅱ): Regionalization of marine fishery. Hangzhou: Zhejiang Science and Technology Press, 1990: 112-234 [《中国渔业资源调查和区划》编辑委员会. 中国渔业资源调查和区划之二中国海洋渔业区划. 杭州: 浙江科学技术出版社, 1990: 112-234]
|
FENG Z Q, HUANG Y W, DONG Z H, et al. Opinions on rational utilization of Acete chinensis resources and fishing closed season of rotating shaft network in the west of Bohai Sea. Marine Fisheries, 1987, 1: 18-20 [冯稚青, 黄燕文, 董志华, 等. 关于合理利用渤海西部毛虾资源和转轴网禁渔期的意见. 海洋渔业, 1987, 1: 18-20] |
FENG Z Q, SUN J H, YANG G B, et al. Discussion of Acete chinensis resources in the western areas of Bohai Sea and their rational use. Transactions of Oceanology and Limnology, 1982, 4: 62-68 [冯志青, 孙介华, 杨贵本, 等. 渤海西部海区毛虾资源现状及合理利用的探讨. 海洋湖沼通报, 1982, 4: 62-68] |
Fisheries Bureau of Ministry of Agriculture, Yellow Sea Fisheries Headquarters of Ministry of Agriculture. Investigation and regionalization of fishery resources in the Yellow Sea and Bohai Sea. Beijing: China Ocean Press, 1990: 141-162 [农业部水产局, 农业部黄海区渔业指挥部. 黄、渤海区渔业资源调查与区划. 北京: 海洋出版社, 1990: 141-162]
|
FREE C M, THORSON J T, PINSKY M L, et al. Impacts of historical warming on marine fisheries production. Science, 2019, 363(6430): 979-983 DOI:10.1126/science.aau1758 |
GONG Y. Influencing actor of nutrient transport in the lower reach of Yellow River. Doctoral Dissertation of Ocean University of China, 2012 [巩瑶. 黄河下游利津站营养盐输送规律及影响因素研究. 中国海洋大学博士研究生学位论文, 2012]
|
GU D X, WANG T, WANG N, et al. Distribution and affecting factors of pseudozoea density of mantis shrimp Oratosquilla oratoria in Bohai Bay. Journal of Dalian Ocean University, 2018, 31(1): 65-71 [谷德贤, 王婷, 王娜, 等. 渤海湾口虾蛄假蚤状幼体的密度分布及影响因素研究. 大连海洋大学学报, 2018, 31(1): 65-71] |
GUAN W J, TIAN S Q, WANG X F, et al. A review of methods and model selection for standardizing CPUE. Journal of Fishery Sciences of China, 2014, 21(4): 852-862 [官文江, 田思泉, 王学昉, 等. CPUE标准化方法与模型选择的回顾与展望. 中国水产科学, 2014, 21(4): 852-862] |
GUAN W J. Application of R program in marine fisheries. Beijing: China Ocean Press, 2015: 224-229 [官文江. R语言在海洋渔业中的应用. 北京: 海洋出版社, 2015: 224-229]
|
HARE J A, RICHARDSON D E. The use of early life stages in stock identification studies. In CADRIN S X, KERR L A, MARIANI S (eds) Stock identification methods: Applications in fishery science (2nd Edition). New York, US: Elsevier, 2013, 329–364
|
HJORT J. Fluctuations in the great fisheries of northern Europe, viewed in the light of biological research. Rapports et Procès-Verbaux des Réunions du Conseil Permanent International Pour L'Exploration de la Mer, 1914, 20: 1-228 |
HOLLIDAY F G T, BLAXTER J H S. The effects of salinity on the developing eggs and larvae of the herring. Journal of the Marine Biological Association UK, 1960, 39: 591-603 DOI:10.1017/S0025315400013564 |
HOUDE E D. Fish life dynamics and recruitment variability. Transactions of the American Fisheries Society, 1987, 2: 17-29 |
HSIEH C H, REISS C, WATSON W, et al. A comparison of long-term trends and variability in populations of larvae of exploited and unexploited fishes in the Southern California region: A community approach. Progress in Oceanography, 2005, 67: 160-185 DOI:10.1016/j.pocean.2005.05.002 |
HSIEH C, REISS C S, HUNTER J R, et al. Fishing elevates variability in the abundance of exploited species. Nature, 2006, 443(7113): 859-862 DOI:10.1038/nature05232 |
JIANG Y W, WAN R J, CHEN R S. Investigation of eggs and larvae of osteichthyes in the Bohai Sea. Marine Fisheries Reseach, 1988, 9: 121-149 [姜言伟, 万瑞景, 陈瑞盛. 渤海硬骨鱼类鱼卵、仔稚鱼调查研究. 海洋水产研究, 1988, 9: 121-149] |
JIANG Y W, WAN R J. Studies on morphology and developmental characters in early stage of Cynoglossus semilavis Guüther in the Bohai Sea. Marine Fisheries Research, 1988, 9: 185-192 [姜言伟, 万瑞景. 渤海半滑舌鳎的生殖习性及产卵生态的研究. 海洋水产研究, 1988, 9: 185-192] |
KONG L B, SUN J R, YANG D L, et al. Comparison research on eggs and larvae of Harengula zunasi Bleeker and Clupanodon punctatus (Temminck & Schlegel) in Bohai Bay. Oceanologia et Limnologia Sinica, 1996, 27(2): 169-178 [孔立波, 孙继仁, 杨东莱, 等. 渤海湾青鳞鱼和斑鰶两种鱼卵及仔鱼的比较研究. 海洋与湖沼, 1996, 27(2): 169-178 DOI:10.3321/j.issn:0029-814X.1996.02.009] |
KOSLOW J A, WRIGHT M. Ichthyoplankton sampling design to monitor marine fish populations and communities. Marine Policy, 2016, 68: 55-64 DOI:10.1016/j.marpol.2016.02.011 |
LASKER R. An egg production method for estimating spawning biomass of pelagic fish: application to the northern anchovy, Engraulis mordax. NOAA Technical Report NMFS, 1985, 36 |
LAUREL B J, HURST T P, COPEMAN L A, et al. The role of temperature on the growth and survival of early and late hatching Pacific cod larvae (Gadus macrocephalus). Journal of Plankton Research, 2008, 30: 1051-1060 DOI:10.1093/plankt/fbn057 |
LAURENCE G C, HOWELL W H. Embryology and influence of temperature and salinity on early development and survival of yellowtail flounder Limundti ferruginecr. Marine Ecology Progress Series, 1981, 6: 11-18 DOI:10.3354/meps006011 |
LEI K, MENG W, ZHENG B H, et al. Variations of water and sediment discharges to the western coast of Bohai Bay and the environmental impacts. Acta Scientiae Circumstantiae, 2007, 27(12): 2052-2059 [雷坤, 孟伟, 郑炳辉, 等. 渤海湾西岸入海径流量和输沙量变化及其环境效应. 环境科学学报, 2007, 27(12): 2052-2059 DOI:10.3321/j.issn:0253-2468.2007.12.020] |
LI Y, REN G Y, WANG Q Y, et al. More extreme marine heatwaves in the China Seas during the global warming hiatus. Environmental Research Letters, 2019, 14(10): 104010 DOI:10.1088/1748-9326/ab28bc |
LI Y, WANG Q Y, LI Q Q, et al. An asymmetric variation of hot and cold SST extremes in the China Seas during the recent warming hiatus period. Scientific Reports, 2021, 11(1): 2014 DOI:10.1038/s41598-020-79854-2 |
Liaoning Marine Fisheries Research Institute, Hebei Marine Fisheries Research Institute. Present situation of Acete chinensis resources and suggestions on future production and utilization of them in the Yellow Sea and Bohai Sea. Yellow Sea Fisheries Research Series, 1961, 12: 16-21 [辽宁省海洋水产研究所, 河北省海洋水产研究所. 黄渤海毛虾资源现状及今后生产利用的意见. 黄海水产研究丛刊, 1961, 12: 16-21] |
LIU J J, ZHANG Y, ZHAI H Y, et al. Distribution characteristics and sources of DOM in Bohai Bay seawater during wet season. China Environmental Science, 2021, 41(10): 4802-4810 [刘晶晶, 张彦, 翟洪艳, 等. 丰水期渤海湾水体中DOM的分布特征及来源. 中国环境科学, 2021, 41(10): 4802-4810 DOI:10.3969/j.issn.1000-6923.2021.10.037] |
LIU J, NING P. Species composition and faunal characteristics of fishes in the Yellow Sea. Biodiversity Science, 2011, 19(6): 764-769 [刘静, 宁平. 黄海鱼类组成、区系特征及历史变迁. 生物多样性, 2011, 19(6): 764-769] |
LIU X S. The research of small yellow croaker (Larimichthys polyactis) geographic race (Investigation Report No.56 of the Yellow Sea Fisheries Research Institute, Ministry of Fisheries, P.R.China). Yellow Sea Fisheries Research Series, 1960, 2: 45-61 [刘效舜. 黄、渤海小黄鱼(Pseudosciaena polyactis)种族问题的初步探讨(中华人民共和国水产部黄海水产研究所调查研究报告第56号). 黄海水产研究丛刊, 1960, 2: 45-61] |
LO N C H, SMITH P E, TAKAHASHI M. Egg, larval and juvenile surveys. In JAKOBSEN T, FOGARTY M J, MEGREY B A, et al (eds) Fish reproductive biology: Implications for assessment and management 2nd edn. West Sussex: Wiley-Blackwell, 2016, 227–251
|
LONG Y, WU X, BI N S, et al. Evolution of active Yellow River (Huanghe) Delta under the water and sediment regulation scheme (WSRS). Marine Geology Frontiers, 2017, 33(3): 7-11 [龙跃, 吴晓, 毕乃双, 等. 黄河调水调沙影响下的现行三角洲叶瓣冲淤演化格局. 海洋地质前沿, 2017, 33(3): 7-11] |
Ministry of Agriculture and Rural Affairs. Report on the state of fishery resources in China inshore waters 2020. Beijing: China Agriculture Press, 2021: 38 [农业农村部. 中国近海渔业资源状况公报2020. 北京: 中国农业出版社, 2021: 38]
|
Municipal Office for Tianjin Bureau of Aquatic Products. The concise report on regionalization of fishery in Tianjin. 1980, 1–33 [天津市水产局区划办公室. 天津市简明渔业区划报告. 1980, 1–33]
|
National Marine Data and Information Service, Ministry of Natural Resources. Blue Book on Marine Climate Change in China (2019–2021). Beijing: Science Press, 2020–2022 [自然资源部国家海洋信息中心. 中国气候变化海洋蓝皮书(2019–2021). 北京: 科学出版社, 2020–2022]
|
NIE H T, TAO J H. Impact of coastal exploitation on the eco-environment of Bohai Bay. Ocean Engineering, 2008, 26(3): 44-50 [聂红涛, 陶建华. 渤海湾海岸带开发对近海水环境影响分析. 海洋工程, 2008, 26(3): 44-50] |
NIELSEN J M, ROGERS L A, BRODEUR, R D, et al. Responses of ichthyoplankton assemblages to the recent marine heatwave and previous climate fluctuations in several Northeast Pacific marine ecosystems. Global Change Biology, 2021, 27: 506-520 DOI:10.1111/gcb.15415 |
PEPIN P, ORR D C, ANDERSON J T. Time to hatch and larval size in relation to temperature and egg size in Atlantic cod (Gadus morhua). Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(Suppl1): 2-10 |
PINKAS L, OLIPHANT M S, IVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters. California Department of Fish and Game, Fish Bulletin, 1971, 152: 1-105 |
PINSKY M L, WORM B, FOGARTY M J, et al. Marine taxa track local climate velocities. Science, 2013, 341(6151): 1239-1242 DOI:10.1126/science.1239352 |
PÖRTNER H O, PECK M A. Climate change effects on fishes and fisheries: Towards a cause-and-effect understanding. Journal of Fish Biology, 2010, 77: 1745-1779 DOI:10.1111/j.1095-8649.2010.02783.x |
QIU D L, LIU X S, WANG Z X. Migration of the small yellow croaker Larimichthys polyactis in the Bohai Sea and Yellow Sea in relation to the hydrographic conditions. In Editorial Committee of the China Scientific Council on the Marine Fisheries Resources 1962 (ed) Anthology sequel of papers on marine fishery resources. Beijing: Agriculture Press, 1965, 43−55 [邱道立, 刘效舜, 王遵孝. 渤、黄海小黄鱼的洄游与水文环境的关系. //1962年海洋渔业资源学术会议论文编审委员会, 海洋渔业资源论文选集续集. 北京: 农业出版社, 1965, 43−55]
|
Research Group on Suitable Fishing Period of Acete chinensis in the Bohai Bay and Laizhou Bay. Preliminary analysis on the composition of catches from set net used for netting Acetes chinensis in Bohai Bay and western Laizhou Bay. Transactions of Oceanology and Limnology, 1992, 4: 76-84 [渤海湾莱州湾毛虾适宜捕捞期的研究课题组. 渤海湾与莱州湾西部毛虾定置网渔获物组成的初步分析. 海洋湖沼通报, 1992, 4: 76-84] |
RIJNSDORP A D, PECK M A, ENGELHARD G H, et al. Resolving the effect of climate change on fish populations. ICES Journal of Marine Science, 2009, 66: 1570-1583 DOI:10.1093/icesjms/fsp056 |
SASSA C, TAKAHASHI M, KONISHI Y, et al. The rapid expansion of yellowtail (Seriola quinqueradiata) spawning ground in the East China Sea is linked to increasing recruitment and spawning stock biomass. ICES Journal of Marine Science, 2019, 77(2): 581-592 |
SAVILLE A, SCHNACK D. Some thoughts on the current status of studies of fish egg and larval distribution and abundance. Rapports et procès-verbaux des réunions-Conseil international pour l'exploration de la mer, 1981, 178: 153-157 |
SHERMAN K, LASKER R, RICHARDS W, et al. Ichthyoplankton and fish recruitment studies in large marine ecosystems. Marine Fisheries Review, 1984, 45: 1-25 |
SOMARAKIS S, TSOUKALI S, GIANNOULAKI M, et al. Spawning stock, egg production and larval survival in relation to small pelagic fish recruitment. Marine Ecology Progress Series, 2019, 617/618: 113-136 DOI:10.3354/meps12642 |
SUN B S, ZUO S H, XIE H L, et al. Analysis of impact effects and changes of the coastline in the Bohai Bay during the past 40 years. Journal of East China Normal University (Natural Science), 2017, 4: 139-148 [孙百顺, 左书华, 谢华亮, 等. 近40年来渤海湾岸线变化及影响分析. 华东师范大学学报(自然科学版), 2017, 4: 139-148 DOI:10.3969/j.issn.1000-5641.2017.04.012] |
SUN X Y, LÜ T T, GAO Y, et al. Driving force analysis of Bohai Bay coastline change from 2000 to 2010. Resources Science, 2014, 36(2): 413-419 [孙晓宇, 吕婷婷, 高义, 等. 2000—2010年渤海湾岸线变迁及驱动力分析. 资源科学, 2014, 36(2): 413-419] |
SUN Z Z, ZHOU J, WANG J, et al. Stow net fishery in the Yellow Sea and Bohai Sea area. Progress in Fishery Sciences, 2012, 33(3): 94-101 [孙中之, 周军, 王俊, 等. 黄渤海区张网渔业. 渔业科学进展, 2012, 33(3): 94-101 DOI:10.3969/j.issn.1000-7075.2012.03.014] |
TAKASUKA A, YONEDA M, OOZEKI Y. Density dependence in total egg production per spawner for marine fish. Fish and Fisheries, 2019, 20: 125-137 DOI:10.1111/faf.12327 |
TANG Q S, YE M Z. Exploitation and protection of offshore fishery resources in Shandong Province. Beijing: China Agriculture Press, 1990: 203-212 [唐启升, 叶懋中. 山东近海渔业资源开发与保护. 北京: 中国农业出版社, 1990: 203-212]
|
THOMPSON A R, BEN-ADERET N J, BOWLIN N M, et al. Putting the Pacific marine heatwave into perspective: The response of larval fish off southern California to unprecedented warming in 2014-2016 relative to the previous 65 years. Global Change Biology, 2021, 28: 1766-1785 |
TIAN M C, SUN B L, YANG J M. Analysis of the fish fauna of the Bohai Sea. Studia Marina Sinica, 1993, 34: 157-167 [田明诚, 孙宝龄, 杨纪明. 渤海鱼类区系分析. 海洋科学集刊, 1993, 34: 157-167] |
WAN R J, CHEN R S. Reproductive behavior and early development of Lateolabrax japonicas (Cuvier et Valenciennes) in the Bohai Sea. Marine Fisheries Research, 1988, 9: 203-211 [万瑞景, 陈瑞盛. 渤海鲈鱼的生殖习性及早期发育特征研究. 海洋水产研究, 1988, 9: 203-211] |
WAN R J, JIANG Y W. The distribution and variation of eggs and larvae of osteichthyes in the Bohai Sea. Journal of Fishery Sciences of China, 1998, 5(1): 43-50 [万瑞景, 姜言伟. 渤海硬骨鱼类鱼卵和仔稚鱼分布及其动态变化. 中国水产科学, 1998, 5(1): 43-50 DOI:10.3321/j.issn:1005-8737.1998.01.008] |
WANG H J, BI N S, SAITO Y, et al. Recent changes in sediment delivery by the Huanghe (Yellow River) to the sea: Causes and environmental implications in its estuary. Journal of Hydrology, 2010, 391(3/4): 302-313 |
WANG X L, CUI Z G, GUO Q, et al. Distribution of nutrients and eutrophication assessment in the Bohai Sea of China. Chinese Journal of Oceanology and Limnology, 2009, 27(1): 177-183 DOI:10.1007/s00343-009-0177-x |
WANG Y Z, WU D, SHI H H, et al. Impact of reclamation on water exchange in Bohai Bay in recent decade. Oceanologia et Limnologia Sinica, 2015, 46(3): 471-480 [王勇智, 吴頔, 石洪华, 等. 近十年来渤海湾围填海工程对渤海湾水交换的影响. 海洋与湖沼, 2015, 46(3): 471-480] |
WEI S. Current situation and prospect of Spanish mackerel resources in Bohai Sea, Yellow Sea and Northern East China Sea (Investigation Report No.184 of the Yellow Sea Fisheries Research Institute, State Aquatic Product General Bureau). Marine Fisheries Research Series, 1981, 27: 9-21 [韦晟. 渤、黄海及东海北部鲅鱼资源的现状和前景(国家水产总局黄海水产研究所调查研究报告第184号). 海洋水产研究丛刊, 1981, 27: 9-21] |
WHITTAKER R H. Evolution and measurement of species diversity. Taxon, 1972, 21(2/3): 213-251 |
WU D X, WAN X Q, BAO X W, et al. Comparison of summer thermohaline field and circulation structure of the Bohai Sea between 1958 and 2000. Chinese Science Bulletin, 2004, 49(3): 287-292 [吴德星, 万修全, 鲍献文, 等. 渤海1958年和2000年夏季温盐场及环流结构的比较. 科学通报, 2004, 49(3): 287-292 DOI:10.3321/j.issn:0023-074X.2004.03.017] |
WU G Z, YANG D L, PANG H Y. The spawning of perch Lateolabrax japonicus (Cuvier & Valenciennes) in Bobai Bay. Marine Science, 1983, 6: 40-45 [吴光宗, 杨东莱, 庞鸿艳. 渤海湾鲈鱼鱼卵和仔、稚鱼分布的研究. 海洋科学, 1983, 6: 40-45] |
WU Z, ZHAO D, SYVITSKI J P M, et al. Anthropogenic impacts on the decreasing sediment loads of nine major rivers in China, 1954-2015. Science of the Total Environment, 2020, 739: 139653 DOI:10.1016/j.scitotenv.2020.139653 |
XIA S F. A preliminary report on the possible impact of the radical cure of Yellow River to the fishery resources in the estuary and adjacent sea areas (Investigation Report No.54 of the Yellow Sea Fisheries Research Institute, Ministry of Fisheries, P.R.China). Yellow Sea Fisheries Research Series, 1960, 2: 18-35 [夏世福. 黄河根治对河口附近海区渔业资源的可能影响初步报告(中华人民共和国水产部黄海水产研究所调查研究报告第54号). 黄海水产研究丛刊, 1960, 2: 18-35] |
XU S S, SONG J M, YUAN H M, et al. Effects of the combined pollution of Cd, Hg, Pb and petroleum hydrocarbons on common fishery species in the Bohai Bay. Asian Journal of Ecotoxicology, 2010, 5(6): 793-802 [许思思, 宋金明, 袁华茂, 等. 镉、汞、铅和石油烃复合污染对渤海湾常见渔业资源生物的影响初探. 生态毒理学报, 2010, 5(6): 793-802] |
YANG D L, WU G Z, PANG H Y. Distribution of eggs and larvae of tongue sole Cynoglossus semilaevis Günther and C. joyneri Günther in Bohai Bay. Chinese Journal of Ecology, 1984(3): 30-33 [杨东莱, 吴光宗, 庞鸿艳. 渤海湾的半滑舌鳎及焦氏舌鳎的鱼卵和仔、稚鱼的季节分布. 生态学杂志, 1984(3): 30-33] |
Yellow River Conservancy Commission of Ministry of Water Resources. Yellow River Sediment Bulletin (2002–2021) [水利部黄河水利委员会. 黄河泥沙公报(2002–2021)]
|
Yellow Sea Fisheries Research Institute, Ministry of Fisheries, P.R.China. Relationship between marine hydrology, meteorology, biological environment and fishery// General situation of fishery resources in the Yellow Sea and Bohai Sea and preliminary suggestions on their development and utilization(Investigation Report No.60 of the Yellow Sea Fisheries Research Institute of the Ministry of Fisheries, P.R.C.). Yellow Sea Fisheries Research Series, 1960, 3: 3−20 [中华人民共和国水产部黄海水产研究所. 海洋水文、气象及生物环境与渔业的关系//黄渤海渔业资源概况及其开发利用的初步意见(中华人民共和国水产部黄海水产研究所调查研究报告第60号). 黄海水产研究丛刊, 1960, 3: 3−20]
|
ZHANG Y X, BIAN X D, SHAN X J, et al. Community structure and suitable habitat for the early life stages of marine fish in the Yantai-Weihai offshore waters. Progress in Fishery Sciences, 2022b, 43(6): 148-167 [张雨轩, 卞晓东, 单秀娟, 等. 烟威近岸海域鱼类早期资源群落结构及适宜产卵生境. 渔业科学进展, 2022b, 43(6): 148-167] |
ZHANG Y X, BIAN X D, SHAN X J, et al. Spatio-temporal distribution of chub mackerel (Scomber japonicus) spawning grounds and its relationship with environmental factors in the offshore waters of Yantai-Weihai. Journal of Fishery Sciences of China, 2022a, 29(4): 618-632 [张雨轩, 卞晓东, 单秀娟, 等. 烟威近岸海域鲐产卵场时空分布及其与环境因子的关系. 中国水产科学, 2022a, 29(4): 618-632] |
ZHAO H P, LI Q X, TAO J H. Spatio-temporal water quality variations and identification of surface water pollutant sources in Bohai Bay. Journal of Hydroelectric Engineering, 2016, 35(10): 21-30 [赵海萍, 李清雪, 陶建华. 渤海湾表层水质时空变化及污染源识别. 水力发电学报, 2016, 35(10): 21-30 DOI:10.11660/slfdxb.20161003] |
ZHAO W, WANG J J, XU S, et al. Distribution characteristics and influencing factors of bacterioplankton community with offshore distance variation in the surface seawater of Bohai Bay. Haiyang Xuebao, 2019, 41(12): 156-171 [赵维, 王敬敬, 徐松, 等. 渤海湾表层海水中浮游细菌群落随离岸距离的分布特征及其影响因素. 海洋学报, 2019, 41(12): 156-171 DOI:10.3969/j.issn.0253-4193.2019.12.015] |
ZHU C J, ZHAO Z C, JI Z Y. Preliminary studies on reproductive characteristics and field forecasting method of small yellow croaker along the coast of Hebei Province. In Editorial Committee of the China Scientific Council on the Marine Fisheries Resources 1962(ed) Anthology sequel of papers on marine fishery resources. Beijing: Agriculture Press, 1965, 84−108 [朱崇俭, 赵增才, 季忠英. 河北沿海小黄鱼的生殖习性及其现场预报方法的初步探讨. //1962年海洋渔业资源学术会议论文编审委员会, 海洋渔业资源论文选集续集. 北京: 农业出版社, 1965, 84−108]
|
ZOU L, XIA J, ZHANG Y, et al. Spatial-temporal change characteristics and driving forces of precipitation in the Haihe River Basin. Water Resources Protection, 2021, 37(1): 53-60 [邹磊, 夏军, 张印, 等. 海河流域降水时空演变特征及其驱动力分析. 水资源保护, 2021, 37(1): 53-60] |