



2. 山东长岛近海渔业资源国家野外科学观测研究站 山东 烟台 265800
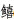 (Sillago japonica)、白姑鱼(Pennahia argentata)、绯
(Sillago japonica)、白姑鱼(Pennahia argentata)、绯 (Callionymus beniteguri)、短吻红舌鳎(Cynoglossus joyneri)等,鱼卵密集区位于调查水域东部至西南部的庙岛海峡毗邻水域;仔稚鱼主要集中于5―7月和11―12月,主要种类为鳀、白氏银汉鱼(Hypoatherina valenciennei)、大泷六线鱼(Hexagrammos otakii)、玉筋鱼(Ammodytes personatus)等,仔稚鱼密集区位于调查水域东部、西南部和西北部;春夏季(5―7月)鱼类早期资源群落功能丰富度、均匀度和离散度指数最高,高值区主要分布在调查水域东北部和西南部。长岛毗邻水域鱼卵及仔稚鱼丰度、群落结构及功能具有明显的时空异质性,鱼类繁殖群体的洄游迁徙对长岛毗邻水域鱼类早期资源群落产生深刻影响。
(Callionymus beniteguri)、短吻红舌鳎(Cynoglossus joyneri)等,鱼卵密集区位于调查水域东部至西南部的庙岛海峡毗邻水域;仔稚鱼主要集中于5―7月和11―12月,主要种类为鳀、白氏银汉鱼(Hypoatherina valenciennei)、大泷六线鱼(Hexagrammos otakii)、玉筋鱼(Ammodytes personatus)等,仔稚鱼密集区位于调查水域东部、西南部和西北部;春夏季(5―7月)鱼类早期资源群落功能丰富度、均匀度和离散度指数最高,高值区主要分布在调查水域东北部和西南部。长岛毗邻水域鱼卵及仔稚鱼丰度、群落结构及功能具有明显的时空异质性,鱼类繁殖群体的洄游迁徙对长岛毗邻水域鱼类早期资源群落产生深刻影响。2. Shandong Changdao National Observation and Research Station for Fishery Resources, Yantai 265800, China
早期生活史阶段是海洋鱼类通过产卵、孵化和发育完成种群补充过程的关键时期,鱼类早期资源丰度及成活率的高低深刻影响地种群补充效率(Hovenkamp, 1992; Dippold et al, 2016),通过科学有效的手段管理鱼类繁殖群体、早期资源及其栖息地生境,对于鱼类种群繁衍及群落稳定性具有关键意义。鱼类早期资源群落功能多样性反映种群的生态位分化水平(Arantes et al, 2019; Mouillot et al, 2007),深入研究关键水域中鱼类早期资源群落结构及功能,可以有效判别鱼类繁殖群体及0龄幼鱼的生境适应程度、资源利用能力及种间竞争强弱,掌握鱼类关键栖息地中不同功能群的生态韧性,对海洋鱼类资源的可持续管理与利用具有重要参考价值。
长岛毗邻水域是黄渤海水体的联通隘口,东临烟威近岸产卵场(张雨轩等, 2022b),西接莱州湾(卞晓东等, 2022)和渤海湾产卵场(万瑞景等, 1998a),北通辽东湾(于旭光等, 2018)和北黄海产卵场(于旭光等, 2020),是众多海洋鱼类的洄游必经之路,也是黄渤海鱼类连通的关键区域,对整个黄渤海渔业资源的补充起着重要作用。长岛毗邻水域鱼类早期资源群落具有黄渤海各产卵场的共性,也具有黄渤海岛群毗邻水域和鱼类洄游过路产卵场的特性。掌握长岛毗邻水域鱼类早期资源群落更替规律及功能多样性,有助于整体把握黄海中北部至渤海大面水域内的鱼类早期生活史,也有助于深入理解温带海岛毗邻水域生态系统中的鱼类种群补充过程,可为黄渤海水域鱼类补充过程提供重要科学依据。目前,针对长岛毗邻水域鱼类早期资源的研究鲜见报道。因此,本研究依托山东长岛近海渔业资源国家野外科学观测研究站,于2021年3―12月在长岛毗邻水域开展鱼卵、仔稚鱼逐月调查,旨在阐释鱼类早期资源丰度时空格局、群落优势种、重要种以及群落分歧种变化,解析鱼类早期资源群落结构更替规律和功能多样性,以期为黄渤海生态廊道长岛毗邻水域鱼类早期生活史特征研究、长岛国家公园建设提供参考。
1 材料与方法 1.1 数据采集与样品分析2021年3―12月,依托于山东长岛近海渔业资源国家野外科学观测研究站,开展了长岛毗邻水域10航次鱼卵、仔稚鱼及生态环境逐月调查,调查站位10个(图 1),调查区域覆盖南五岛(大黑山岛、小黑山岛、庙岛、北长山岛和南长山岛)近岸水域,调查日期如表 1所示。调查船为鲁昌渔64756及鲁昌渔65678 (调查船参数:船长23.24 m,型宽5 m,型深2.15 m,主机总功率110 kW,总吨位65 t),海上调查严格按照《海洋渔业资源调查规范》(SC/T 9403-2012)。

|
图 1 长岛毗邻水域鱼类早期资源采样区域及站位 Fig.1 Sampling areas and stations for early life fish resources in the waters adjacent to Changdao Islands |
|
|
表 1 2021年长岛毗邻水域鱼类早期资源调查时间 Tab.1 Survey dates of early life fish resources in the waters adjacent to Changdao Islands in 2021 |
使用标准大型浮游生物网(网口内径0.8 m,网目孔径0.505 mm)于海水表层迎流水平拖网10 min,拖速3 n mile/h,采集鱼卵、仔稚鱼样品,样品通过5%海水甲醛溶液固定保存,实验室内进行全样本物种鉴定,确认至“种”单元,并按照种类、存活状态和发育阶段分别记录个体数。鉴定按照《海洋渔业资源调查规范》(SC/T 9403-2012)。物种拉丁文名引自World Register of Marine Species (WoRMS, https://www.marinespecies.org/),形态学鉴定参考张仁斋等(1985)、冲山宗雄(2014)和万瑞景等(1998a、1998b、2016)。水温数据通过德国Sea & Sun Technology CTD 60M温盐深仪匀速垂直采集,取0~0.5 m水深区间内的水温平均值作为海水表层温度(sea surface temperature, SST)。
1.2 资源丰度时空格局通过单位捕捞努力量渔获量(catch per unit effort, CPUE)对鱼卵及仔稚鱼的丰度水平进行量化,以每10 min水平拖网作业捕获的鱼卵或仔稚鱼个体数作为CPUE (ind./10 min)。运用Garrison分布重心法(Garrison, 2001)计算鱼卵、仔稚鱼的CPUE重心,以表示鱼类早期资源分布重心迁移。CPUE重心坐标的计算公式如下:
| $ X = \frac{{\sum\limits_{i = 1}^n {{X_i} \times {N_i}} }}{{\sum\limits_{i = 1}^n {{N_i}} }} $ | (1) |
| $ Y = \frac{{\sum\limits_{i = 1}^n {{Y_i} \times {N_i}} }}{{\sum\limits_{i = 1}^n {{N_i}} }} $ | (2) |
式中,X为CPUE重心的经度,Y为CPUE重心的纬度,Xi为i站位的经度,Yi为i站位的纬度,Ni为i站位鱼类早期资源个体数(ind.)。
1.3 物种优势度通过Pinkas相对重要性指数(index of relative importance, IRI) (Pinkas, 1971)判断2021年3―12月各月鱼卵及仔稚鱼的优势种(IRI≥1 000)、重要种(1 000 > IRI≥100)、常见种(100 > IRI > 10)、一般种(10 > IRI > 1)和少见种(IRI < 1) (程济生, 2004),计算公式如下:
| $ {\text{IRI}} = N\% \times F\% \times {10^4} $ | (3) |
式中,N%为某种鱼卵或仔稚鱼的数量占当月鱼卵或仔稚鱼总数量的百分比,F%为某种鱼卵或仔稚鱼的站位出现频率。
1.4 群落结构月际更替以物种更替率作为2021年3―12月各相邻月间长岛毗邻水域鱼类早期资源群落更替幅度的衡量指标,鱼卵及仔稚鱼物种更替率(E)计算公式为(张荣良等, 2021):
| $ E = \frac{A}{{A + B}} $ | (4) |
式中,A为相邻月份之间种类增加数与减少数之和;B为相邻月份之间相同种类数。
相似性百分比分析(similarity percentage analysis, SIMPER)可基于Bray-Curtis距离评估物种对群落组内相似度和组间相异度的贡献,是分析生态群落特征及更替模式的有效手段(Pinheiro et al, 2016; Drakard et al, 2018; 栾青杉等, 2018)。将各月鱼类早期资源种群CPUE矩阵进行平方根转换(李建都等, 2023)以削弱高CPUE种群的相异度贡献率,使低CPUE种群纳入分析过程,以各月各种群CPUE平方根构建二维矩阵,对鱼卵、仔稚鱼群落进行相邻月间SIMPER分析,将各种群按照平均相异度贡献率百分比(contribution percentage of average dissimilarities, C)由高至低排序,当累积贡献率百分比达90% (芮银等, 2022)时停止排序,以加入排序的种群集合为相邻月间鱼类早期资源群落的主要分歧种。交替月份组内分歧种的相对CPUE差值可反映单一种群分布的月份偏好,计算公式为:
| $ {\text{CPUE}}_{ri}=\frac{{\text{CPUE}}_{i}}{{\text{CPUE}}_{ tot}}\times 100\% $ | (5) |
| $ {\text{CPU}}{{\text{E}}_{rdi}} = {\text{CPU}}{{\text{E}}_{ri}} - {\text{CPU}}{{\text{E}}_{r(i - 1)}} $ | (6) |
式中,CPUEi为某分歧种在i月的CPUE,CPUEtot为该分歧种各月CPUE总和,CPUEri为该分歧种在i月的相对CPUE,CPUEr(i–1)为该分歧种在(i–1)月的相对CPUE,CPUErdi为i月该分歧种的相对CPUE较上个月的差值。
1.5 群落功能特征指数海洋鱼类的适温类型和栖息地类型深刻影响鱼类繁殖群体的时空格局(Yasuda et al, 2023; Sabatés et al, 2007),而鱼类产卵类型影响了鱼卵及初孵仔鱼的分布方位及水层(Baras et al, 2018; Tarimo et al, 2022),因此,基于鱼类群落中各种群适温类型、栖息地类型和产卵类型的性状分布情况,可以表征鱼类种群的生态位分化效果,进而探究不同时期和不同水域内鱼类群落的稳定性。因此,选择以上3个性状作为长岛毗邻水域鱼类早期资源及其繁殖群体群落功能特征的判断依据,共计14个类别(表 2)。基于海表温差异划分4个季节,3月和4月为春季,5月、6月和7月为春夏季,8月和9月为夏秋季,10月、11月和12月为秋冬季,构建各季节鱼类早期资源群落的种群站位分布矩阵;鉴于各鱼类种群存在繁殖力差异,以二分类数据表征种群在各站位出现与否,即各出现种群的丰度权重相等,聚焦鱼类繁殖群体的功能特征;基于相似性分析(analysis of similarities, ANOSIM)验证鱼类早期资源群落的季节差异显著性。基于各季节鱼类早期资源种群在成鱼期的适温类型、栖息地类型和产卵类型,构建种群性状矩阵,以分类数据表征不同性状类别。基于种群站位分布矩阵和种群性状矩阵,计算各季节各站位鱼类早期资源群落功能丰富度指数(functional richness index, FRic)、均匀度指数(functional evenness index, FEve)和离散度指数(functional divergence index, FDiv),探究鱼类早期资源群落及繁殖群体功能特征的时空差异,FRic、FEve和FDiv计算方法见Villéger等(2008)。
|
|
表 2 鱼类早期资源种群成鱼期功能性状及类别划分 Tab.2 Functional traits and categories of early life fish populations during the adult stage |
SIMPER及ANOSIM基于R语言vegan包进行分析;群落功能特征指数基于R语言FD包进行计算,功能体积计算基于快速凸包算法(Barber et al, 1996),距离矩阵基于Gower距离(Gower, 1971);基于反距离权重插值法(Watson et al, 1985),使用ArcMap 10.2绘制鱼类早期资源CPUE及群落特征指数时空分布图。
2 结果 2.1 鱼卵、仔稚鱼时空分布2021年3―12月鱼卵CPUE分布见图 2。共采集鱼卵71 695粒,其中,3、4、9、11和12月未采集到鱼卵,5月采集鱼卵8 951粒,出现频率为100%,平均CPUE为895.1 ind./10 min,集中分布于南五岛西南、东北部水域,以南五岛西南部CD02站CPUE最高,为5 209 ind./10 min;6月采集鱼卵59 049粒,出现频率为100%,平均CPUE为5 904.9 ind./10 min,除小黑山岛东部CD07站外,其余站位平均CPUE均高于1 000 ind./10 min,以庙岛南部CD04站CPUE最高,为30 199 ind./10 min;7月采集鱼卵3 396粒,出现频率为100%,平均CPUE为339.6 ind./10 min,集中分布于南五岛东、南部水域,北长山岛东北部CD09站CPUE最高,为868 ind./10 min;8月采集鱼卵298粒,出现频率为90%,平均CPUE为29.8 ind./10 min,南五岛西、南部水域鱼卵CPUE较高,以南五岛南部CD01站CPUE最高,为181 ind./10 min;10月采集鱼卵1粒,分布于大黑山岛西北部CD08站。

|
图 2 长岛毗邻水域鱼卵CPUE及海表温度时空格局 Fig.2 Spatio-temporal patterns of CPUE of fish eggs and sea surface temperature in the waters adjacent to Changdao Islands a:3月;b:4月;c:5月;d:6月;e:7月;f:8月;g:9月;h:10月;i:11月;j:12月 a: March; b: April; c: May; d: June; e: July; f: August; g: September; h: October; i: November; j: December |
2021年3―12月仔稚鱼CPUE分布如图 3所示。共采集仔稚鱼1 161尾,其中,3月采集仔稚鱼10尾,出现频率为20%,平均CPUE为1 ind./10 min,以南五岛北部CD10站CPUE最高,为8 ind./10 min;4月采集仔稚鱼27尾,出现频率为50%,平均CPUE为2.7 ind./10 min,集中分布于南五岛中南部,以南部CD01站CPUE最高,为14 ind./10 min;5月采集仔稚鱼446尾,出现频率为60%,平均CPUE为44.6 ind./10 min,集中分布于南、北长山岛以西,以南五岛西南部CD02站CPUE最高,为392 ind./10 min;6月采集仔稚鱼48尾,出现频率为60%,平均CPUE为4.8 ind./10 min,集中分布于南五岛中西部,以大黑山岛西部CD05站CPUE最高,为16 ind./10 min;7月采集仔稚鱼159尾,出现频率为80%,平均CPUE为15.9 ind./10 min,集中分布于南五岛西南部、东部,以南长山岛东部CD06站CPUE最高,为91 ind./10 min;8月采集仔稚鱼14尾,出现频率为60%,平均CPUE为1.4 ind./10 min,以大黑山岛西部CD05站CPUE最高,为5 ind./10 min;9月采集仔稚鱼3尾,出现频率为20%,平均CPUE为0.3 ind./10 min,分布于南五岛南部CD01及西部CD05站;10月采集仔稚鱼32尾,出现频率为70%,平均CPUE为3.2 ind./10 min,分布于南五岛西南部、东部,南五岛西南部CD02站CPUE最高,为14 ind./10 min;11月采集仔稚鱼241尾,出现频率为90%,平均CPUE为24.1 ind./10 min,除南五岛西南部、东南部外,其余大面水域仔稚鱼CPUE均较高,以南五岛北部CD10站CPUE最高,为77 ind./10 min;12月采集仔稚鱼181尾,出现频率为100%,平均CPUE为18.1 ind./10 min,除南五岛西南部、中部外,其余水域仔稚鱼CPUE较高,以南长山岛东部CD06站CPUE最高,为45 ind./10 min。

|
图 3 长岛毗邻水域仔稚鱼CPUE及海表温度时空格局 Fig.3 Spatio-temporal patterns of CPUE of fish larvae and sea surface temperature in the waters adjacent to Changdao Islands a: 3月; b: 4月; c: 5月; d: 6月; e: 7月; f: 8月; g: 9月; h: 10月; i: 11月; j: 12月。 a: March; b: April; c: May; d: June; e: July; f: August; g: September; h: October; i: November; j: December. |
2021年3―12月鱼类早期资源CPUE重心月际迁移状况见图 4。在主要产卵期5―8月,鱼卵CPUE重心由大黑山岛南部近岸水域东移至庙岛南部近岸水域,而后继续东移至南长山岛北部水域,最后迁移至庙岛南部水域。在仔稚鱼主要分布期内,5―7月仔稚鱼CPUE重心自南五岛西南部移至小黑山岛南部水域,而后向东南迁移至南长山岛西北部近岸水域;11―12月仔稚鱼CPUE重心自小黑山岛东北部近岸水域移至庙岛东北部近岸水域。

|
图 4 长岛毗邻水域鱼类早期资源CPUE重心的月际迁移 Fig.4 Distribution patterns of the CPUE centroids of early life fish resources in the waters adjacent to Changdao Islands a:5月、6月、7月、8月和10月鱼卵CPUE重心;b:3―12月仔稚鱼CPUE重心。 a: CPUE centroids of fish eggs in May, June, July, August, and October; b: CPUE centroids of fish larvae from March to December. |
2021年3―12月共采集21种鱼卵,隶属6目16科,鲈形目(Perciformes)含9科13种,鲱形目(Clupeiformes)含2科3种,鲽形目(Pleuronectiformes)含2科2种,灯笼鱼目(Myctophiformes)、鲻形目(Mugiliformes)和

|
|
表 3 2021年3―12月长岛毗邻水域鱼类早期资源种类 Tab.3 Species of early life fish resources in the waters adjacent to Changdao Islands from March to December, in 2021 |
 Konosirus punctatus
Konosirus punctatus 科Hemiramphidae
科Hemiramphidae Hyporhamphus sajori
Hyporhamphus sajori Planiliza haematocheilus
Planiliza haematocheilus 科Sillaginidae
科Sillaginidae Sillago japonica
Sillago japonica Seriola lalandi
Seriola lalandi 科Callionymidae
科Callionymidae Callionymus beniteguri
Callionymus beniteguri Callionymus valenciennei
Callionymus valenciennei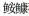 科Lophiidae
科Lophiidae Lophius litulon
Lophius litulon2021年3―12月鱼类早期资源种类数及适温类型见图 5。3月,共采集2种冷温性仔稚鱼;4月,共采集3种冷温性仔稚鱼;5月,共采集11种鱼类早期资源,9种暖温性和1种暖水性鱼卵,1种冷温性和2种暖温性仔稚鱼;6月,共采集20种鱼类早期资源,7种暖温性和5种暖水性鱼卵,1种冷温性和8种暖温性仔稚鱼;7月,共采集16种鱼类早期资源,5种暖温性和5种暖水性鱼卵,5种暖温性和3种暖水性仔稚鱼;8月,共采集9种鱼类早期资源,3种暖温性和3种暖水性鱼卵,1种暖温性和2种暖水性仔稚鱼;9月,共采集1种暖温性仔稚鱼;10月,共采集1种暖温性鱼卵,1种冷温性和1种暖温性仔稚鱼;11月,共采集1种冷温性和2种暖温性仔稚鱼;12月,共采集4种冷温性仔稚鱼。

|
图 5 长岛毗邻水域鱼类早期资源种类数及其成鱼期适温类型
Fig.5 Number of species of early life fish resources and their temperature adaptation types during the adult stage in the waters adjacent to Changdao Islands
 :鱼卵、仔稚鱼种类总数; :鱼卵、仔稚鱼种类总数; :鱼卵暖温种; :鱼卵暖温种; :鱼卵暖水种; :鱼卵暖水种; :仔稚鱼冷温种; :仔稚鱼冷温种; :仔稚鱼暖温种; :仔稚鱼暖温种; :仔稚鱼暖水种。
: Total number of species of fish eggs and larvae; : Warm-temperate fish egg species; : Warm-water fish egg species; : Cold-temperate fish larva species; : Warm-temperate fish larva species; : Warm-water fish larva species. :仔稚鱼暖水种。
: Total number of species of fish eggs and larvae; : Warm-temperate fish egg species; : Warm-water fish egg species; : Cold-temperate fish larva species; : Warm-temperate fish larva species; : Warm-water fish larva species.
|
2021年3―12月长岛毗邻水域鱼类早期资源优势种与重要种的月际变动如表 4所示。3月仔稚鱼优势种为大泷六线鱼,重要种为玉筋鱼(Ammodytes personatus)。4月仔稚鱼优势种为方氏云鳚(Pholis fangi)和大泷六线鱼,无重要种。5月鱼卵优势种为鳀,无重要种;仔稚鱼优势种为鳀,无重要种。6月鱼卵优势种为鳀,无重要种;仔稚鱼无优势种,重要种为普氏细棘
|
|
表 4 长岛毗邻水域鱼类早期资源优势种与重要种 Tab.4 Dominant and important species of early life fish resources in the waters adjacent to Changdao Islands |
 H. sajori
H. sajori H. sajori
H. sajori C. beniteguri
C. beniteguri2021年3―12月鱼类早期资源物种更替率如图 6所示。鱼卵物种更替率在4―8月期间由100%逐月递减40%,9月、10月均升至100%,仔稚鱼物种更替率在3―5月由33.33%升至100%,5―7月递减至69.23%,7―9月逐月回升至100%并维持至10月,11月骤降至33.33%,11―12月升高至83.33%。综合看来,4―5月、8―9月和9―10月为鱼卵和仔稚鱼群落的完全更替期,7―8月鱼卵种类组成相似度最高,3―4月和10―11月仔稚鱼群落的种群组成相似度最高;除5―8月外,其他月份囿于较少的鱼类早期资源种类,邻月群落的物种更替率波动幅度较大,5―8月鱼卵物种更替率逐渐降至较低水平(40%),邻月群落的种类组成趋于相似,仔稚鱼群落的物种更替率曲线位于鱼卵群落上方,处于较高水平(E > 60%),与鱼卵群落相比,仔稚鱼群落月际更替幅度更大。

|
图 6 长岛毗邻水域鱼类早期资源群落物种更替率 Fig.6 Monthly turnover rates of early life fish communities in the waters adjacent to Changdao Islands |
2021年3―12月长岛毗邻水域鱼类早期资源群落月际更替的主要分歧种及相对CPUE如表 5所示。3―4月,仔稚鱼群落分歧种为方氏云鳚,CPUE大幅增加。4―5月,鳀、黑鲷(Acanthopagrus schlegelii)、蓝点马鲛(Scomberomorus niphonius)、绯
|
|
表 5 月交替期间鱼类早期资源群落分歧种的平均相异性贡献率百分比及相对CPUE差值 Tab.5 Average dissimilarity contribution percentage and relative CPUE differences of divergent species in early life fish communities over the alternation of months |
 C. beniteguri
C. beniteguri K. punctatus
K. punctatus H. sajori
H. sajori P. haematocheilus
P. haematocheilus C. beniteguri
C. beniteguri 虎鱼A. pflaumii
虎鱼A. pflaumii C. beniteguri
C. beniteguri K. punctatus
K. punctatusANOSIM检验结果显示,2021年鱼类早期资源群落季节差异显著(P < 0.01),各季节月份组划分较为合理。站位–种群矩阵以种群出现与否作为判断标准,各出现种群在功能特征指数计算中的权重相等,以重点关注各站位群落的功能性状覆盖状况。2021年春季出现的大泷六线鱼、玉筋鱼和方氏云鳚性状特征相同,故群落功能丰富度、均匀度和离散度均不存在,春夏季、夏秋季和秋冬季鱼类早期资源群落功能特征指数的时空变动情况如图 7所示。时间上看,春夏季鱼类早期资源群落功能丰富度、均匀度与离散度平均值最高,夏秋季次高,秋冬季次低,春季最低。空间上看,春夏季南五岛西南、西北和东北部水域鱼类早期资源群落功能丰富度水平较高,西北部水域群落功能均匀度水平较高,北部和东南部水域群落功能离散度水平较高;夏秋季南五岛西部和南部水域群落功能丰富度水平较高,西部、南部和东北部水域群落功能均匀度和功能离散度水平均较高;秋冬季,南五岛南部水域群落功能丰富度、功能均匀度和功能离散度水平较高。综合来看,长岛毗邻水域鱼类早期资源群落功能丰富度、均匀度和离散度时空差异明显,在有鱼类早期资源分布的水域中,功能离散度均处于较高水平。

|
图 7 长岛毗邻水域鱼类早期资源群落功能特征指数时空分布 Fig.7 Spatio-temporal distributions of the functional trait indices of early life fish communities in the waters adjacent to Changdao Islands a、b、c分别为春夏季、夏秋季和秋冬季的FRic;d、e、f分别为春夏季、夏秋季和秋冬季FEve;g、h、i分别为春夏季、夏秋季和秋冬季FDiv。 a, b, and c: FRic in spring & summer (SS), summer & autumn (SA) and autumn & winter (AW), respectively; d, e, and f: FEve in SS, SA and AW, respectively; g, h, and I: FDiv in SS, SA and AW, respectively. |
长岛毗邻水域鱼类早期资源丰度与种类有明显的时空异质性,水温变动是鱼类早期资源群落更替的重要原因。长岛毗邻水域在春季初期至中期(3―4月)水温普遍较低,中期开始出现西南–东北方向的水温梯度,平均海表温低于9 ℃,大泷六线鱼、玉筋鱼、方氏云鳚等冷温性仔稚鱼集中分布在南五岛中部经向断面的低温水域内。春末夏初(5―6月),长岛毗邻水域进入快速增温期,鳀、黄姑鱼、白姑鱼、长蛇鲻、黑鲷、蓝点马鲛等洄游性鱼类(赵传 , 1990; 陈大刚, 1991)的繁殖群体进入黄海中北部至渤海水域中各大产卵场并开始大规模产卵,作为黄渤海鱼类重要洄游通道,长岛毗邻水域暖温性、暖水性鱼类早期资源种类数与丰度显著上升,达到了调查期间最高水平;与周边产卵场(卞晓东等, 2022; 于旭光等, 2018、2020; 张雨轩等, 2022b)相同,鳀是长岛毗邻水域增温期内鱼卵和仔稚鱼的最优势种类,除南五岛内部浅水区以外,调查水域内鳀卵丰度均较高,密集区位于南五岛西南部至南部水域,该水域较高的表层温度(16~23 ℃)为鳀的大规模产卵提供了适宜水温环境(Funamoto et al, 2004; Takasuka et al, 2007);长岛毗邻水域在夏季初期的水温范围与黄海黄姑鱼产卵场相近(雷霁霖等, 1981),适宜其大规模产卵,黄姑鱼卵丰度在夏初鱼卵群落中仅次于鳀,不过其鱼卵的站位出现频率和分布均匀度均较低,高度集中于南长山岛东部的黄海水域内;蓝点马鲛在近年来成为渤海莱州湾产卵场增温期优势产卵种类(卞晓东等, 2022),本研究在长岛毗邻水域发现其产卵洄游迹象,鱼卵集中分布于调查水域南部和西南部水域。夏季中期至末期(7―8月),长岛毗邻水域水温缓速上升至全年最高(丁东, 1994),海表温水平梯度差异减小,鳀、黄姑鱼、长蛇鲻等在增温期内大规模产卵的暖温性鱼种结束产卵盛期,调查水域鱼卵总丰度大幅下降;少鳞, 1990),在南五岛西北部渤海水域内发现其产卵迹象,但长岛毗邻水域以岩石底质为主(丁东, 1994),与花鲈中心产卵场的细粉砂底、粉砂质粘土软泥底和粗粉砂底(赵传, 1990)存在明显差异,较不适宜花鲈产卵,其鱼卵丰度处于极低水平;已结束索饵的0龄鳀稚鱼零星分布于西南部渤海水域,即将洄游出渤海南下越冬(赵传, 1990);冷温性的大泷六线鱼仔稚鱼出现,主要分布于南五岛南部纬向断面水域;降温期内,洄游性鱼类繁殖群体基本结束产卵洄游活动,鱼类早期资源种类开始向冷温种更替,作为洄游通道的长岛毗邻水域已不存在集中分布的鱼类早期资源,鱼卵、仔稚鱼群落接近断层。秋末冬初(11―12月),长岛毗邻水域进入低温期,水温大幅下降至春季中期水平,大泷六线鱼、玉筋鱼、方氏云鳚、日本笠鳚(Chirolophis japonicus)和花鲈仔稚鱼出现,以大泷六线鱼和玉筋鱼为优势种,其中,大泷六线鱼仔稚鱼暴发式增长,密集区位于南五岛中部、北部和西北部水域,考虑到大泷六线鱼仔稚鱼的胚胎发育期较长(潘雷等, 2012),其在长岛毗邻水域的产卵盛期应为11月初;至此,长岛毗邻水域鱼类早期资源群落更替为冷温性特征。
, 1990; 陈大刚, 1991)的繁殖群体进入黄海中北部至渤海水域中各大产卵场并开始大规模产卵,作为黄渤海鱼类重要洄游通道,长岛毗邻水域暖温性、暖水性鱼类早期资源种类数与丰度显著上升,达到了调查期间最高水平;与周边产卵场(卞晓东等, 2022; 于旭光等, 2018、2020; 张雨轩等, 2022b)相同,鳀是长岛毗邻水域增温期内鱼卵和仔稚鱼的最优势种类,除南五岛内部浅水区以外,调查水域内鳀卵丰度均较高,密集区位于南五岛西南部至南部水域,该水域较高的表层温度(16~23 ℃)为鳀的大规模产卵提供了适宜水温环境(Funamoto et al, 2004; Takasuka et al, 2007);长岛毗邻水域在夏季初期的水温范围与黄海黄姑鱼产卵场相近(雷霁霖等, 1981),适宜其大规模产卵,黄姑鱼卵丰度在夏初鱼卵群落中仅次于鳀,不过其鱼卵的站位出现频率和分布均匀度均较低,高度集中于南长山岛东部的黄海水域内;蓝点马鲛在近年来成为渤海莱州湾产卵场增温期优势产卵种类(卞晓东等, 2022),本研究在长岛毗邻水域发现其产卵洄游迹象,鱼卵集中分布于调查水域南部和西南部水域。夏季中期至末期(7―8月),长岛毗邻水域水温缓速上升至全年最高(丁东, 1994),海表温水平梯度差异减小,鳀、黄姑鱼、长蛇鲻等在增温期内大规模产卵的暖温性鱼种结束产卵盛期,调查水域鱼卵总丰度大幅下降;少鳞, 1990),在南五岛西北部渤海水域内发现其产卵迹象,但长岛毗邻水域以岩石底质为主(丁东, 1994),与花鲈中心产卵场的细粉砂底、粉砂质粘土软泥底和粗粉砂底(赵传, 1990)存在明显差异,较不适宜花鲈产卵,其鱼卵丰度处于极低水平;已结束索饵的0龄鳀稚鱼零星分布于西南部渤海水域,即将洄游出渤海南下越冬(赵传, 1990);冷温性的大泷六线鱼仔稚鱼出现,主要分布于南五岛南部纬向断面水域;降温期内,洄游性鱼类繁殖群体基本结束产卵洄游活动,鱼类早期资源种类开始向冷温种更替,作为洄游通道的长岛毗邻水域已不存在集中分布的鱼类早期资源,鱼卵、仔稚鱼群落接近断层。秋末冬初(11―12月),长岛毗邻水域进入低温期,水温大幅下降至春季中期水平,大泷六线鱼、玉筋鱼、方氏云鳚、日本笠鳚(Chirolophis japonicus)和花鲈仔稚鱼出现,以大泷六线鱼和玉筋鱼为优势种,其中,大泷六线鱼仔稚鱼暴发式增长,密集区位于南五岛中部、北部和西北部水域,考虑到大泷六线鱼仔稚鱼的胚胎发育期较长(潘雷等, 2012),其在长岛毗邻水域的产卵盛期应为11月初;至此,长岛毗邻水域鱼类早期资源群落更替为冷温性特征。
综合来看,2021年长岛毗邻水域鱼类主要产卵期为5―8月增温期,密集区位于南五岛东部至西南部的庙岛海峡毗邻水域,其中,5―6月是鳀、黄姑鱼、长蛇鲻、黑鲷等暖温性鱼类的产卵盛期,7―8月为绯
生物群落的性状特征对生态系统功能产生重要影响(Hooper et al, 2004; Bellwood et al, 2004),群落功能多样性是生物多样性的重要组成部分(Díaz et al, 2001),基于群落性状丰富度、均匀度、离散度等指标,可以量化群落性状范围及分布特征,掌握群落功能多样性的时空格局(Villéger et al, 2008; 邹建宇等, 2023; 张晓妆等, 2019)。长岛毗邻水域鱼类早期资源群落功能特征的时空差异明显。囿于较低的水温,春季鱼类早期资源种类仅包含大泷六线鱼、玉筋鱼和方氏云鳚,在成鱼期均为冷温性、大陆架浅水底层、产粘着沉性卵鱼类,群落功能多样性较差。在海水升温明显的春夏季,鱼类大规模迁入黄渤海产卵洄游,增加了长岛毗邻水域鱼类繁殖群体种类,使鱼类早期资源群落占据更多的生态位空间,优势适温类型、栖地类型和产卵类型分别为暖温性、大陆架浅水底层及中底层、产漂流浮性卵,群落功能多样性为调查期间最高水平;南五岛东南部水域鱼类早期资源种类丰富,但群落功能均匀度较低,栖息于大陆架浅水中上层和底层且产浮性卵的暖温性种群冗余度较高,生态位分化效果不佳,新生的0龄幼鱼群落或将存在较强的种间竞争作用;西北部水域鱼类早期资源种类数较少,群落功能丰富度和离散度极高,冗余度较低,种间作用较弱,由此水域向邻近产卵场输送的鱼类繁殖群体具有较高的功能多样性,但局部群落中单个种群受环境和人类活动负面影响造成的功能性状缺口不易及时得到补全;东北部和西南部水域鱼类早期资源种类丰富,且具有较高的群落功能丰富度和离散度以及中等水平的均匀度,局部群落具较强的抗干扰能力,且向邻近产卵场输送的鱼类繁殖群体可占据较大的生态位空间,有利于提高邻近产卵场鱼类早期资源群落稳定性。夏秋季长岛毗邻水域鱼类早期资源群落在成鱼期的优势性状分别为暖水性、大陆架浅水底层、漂流浮性卵,秋冬季群落在成鱼期的优势性状分别为冷温性、大陆架浅水底层、产粘着沉性卵;长岛毗邻水域洄游性鱼类产卵盛期的结束使鱼类早期资源种类数下降,各种群占据的生态位空间缩小,群落功能多样性降低。综合来看,长岛毗邻水域鱼类早期资源群落在春夏季的功能多样性水平最高,其中,南五岛东北部和西南部水域具有最多的鱼类早期资源种类,并且群落功能丰富,性状分布均匀,生态位分化良好,是黄海中北部至渤海各产卵场鱼类繁殖群体及早期资源群落功能补充的关键水域。此外,各季节鱼类早期资源群落功能多样性均呈现内部水域低、周边水域高的分布特征,内部水域的浅水特征(赵永松等, 2022)限制了栖息地性状广度,加之人类活动在行为学和生态学上对鱼类繁殖群体造成双重干扰,导致该水域鱼类早期资源群落功能多样性始终保持极低水平。
栖息地生境的“过滤”作用和种间竞争的“分化”作用共同决定了群落中种群特征(Mayfield et al, 2005),海洋鱼类早期资源群落结构是鱼类繁殖群体对种间竞争作用和栖息地环境的适应性结果。在春夏季增温期,黄海中北部产卵场的优良生境吸引大量鱼类繁殖群体集聚(赵传, 1990),鱼类生态位重叠效应不断增强,产卵场容纳量(唐启升, 1996)逐渐趋向饱和,繁殖群体穿越长岛毗邻水域,在渤海至黄海中北部大面水域内寻觅更适宜的产卵场所,以使后代被捕食概率最小化、食物摄入量最大化(Ciannelli et al, 2015)。作为黄渤海鱼类洄游的必经之路,长岛毗邻水域洄游性鱼类早期资源的丰度及功能多样性水平具有极高的研究价值。因此,对该水域鱼类早期生活史调查数据进行挖掘和分析,未来不应局限于传统产卵场视角,作为鱼类洄游的重要生态廊道,长岛毗邻水域在黄渤海鱼类洄游过程中的连通性同样值得重点关注。
ARANTES C C, FITZGERALD D B, HOEINGHAUS D J, et al. Impacts of hydroelectric dams on fishes and fisheries in tropical rivers through the lens of functional traits. Current Opinion in Environmental Sustainability, 2019, 37: 28-40 DOI:10.1016/j.cosust.2019.04.009 |
BARAS E, ARIFIN O Z, SLEMBROUCK J, et al. Oil globule size in fish eggs: A matter of biome and reproductive strategy. Fish and Fisheries, 2018, 19(6): 996-1002 DOI:10.1111/faf.12307 |
BARBER C B, DOBKIN D P, HUHDANPAA H. The quickhull algorithm for convex hulls. ACM Transactions on Mathematical Software, 1996, 22(4): 469-483 DOI:10.1145/235815.235821 |
BELLWOOD D R, HUGHES T P, FOLKE C, et al. Confronting the coral reef crisis. Nature, 2004, 429(6994): 827-833 DOI:10.1038/nature02691 |
BIAN X D, WAN R J, JIN X S, et al. Early life resources assemblage structure and succession to the marine Osteichthyes in the Laizhou Bay of Bohai Sea. Journal of Fisheries of China, 2022, 46(1): 51-72 [卞晓东, 万瑞景, 金显仕, 等. 渤海莱州湾硬骨鱼类早期资源群落结构及演变. 水产学报, 2022, 46(1): 51-72] |
CHEN D G. Fishery ecology in the Yellow Sea and Bohai Sea. Beijing: Ocean Press, 1991 [陈大刚. 黄渤海渔业生态学. 北京: 海洋出版社, 1991]
|
CHENG J S. Ecological environment and biotic community in the coastal waters of the Yellow Sea and Bohai Sea. Qingdao: China Ocean University Press, 2004, 244-260 [程济生. 黄渤海近岸水域生态环境与生物群落. 青岛: 中国海洋大学出版社, 2004, 244-260] |
CIANNELLI L, BAILEY K, OLSEN E M. Evolutionary and ecological constraints of fish spawning habitats. ICES Journal of Marine Science, 2015, 72(2): 285-296 DOI:10.1093/icesjms/fsu145 |
DÍAZ S, CABIDO M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends in Ecology and Evolution, 2001, 16(11): 646-655 DOI:10.1016/S0169-5347(01)02283-2 |
DING D. Natural conditions of Bohai strait cross sea passage. Marine Geology Letters, 1994(2): 4-6 [丁东. 渤海海峡跨海通道的自然条件. 海洋地质动态, 1994(2): 4-6] |
DIPPOLD D A, LEAF R T, PETERSON M S. Evaluating management regimes using Per-Recruit Models and relative stock density for Mississippi's spotted seatrout. North American Journal of Fisheries Management, 2016, 36(5): 1178-1189 DOI:10.1080/02755947.2016.1204387 |
DRAKARD V F, LANFRANCO S, SCHEMBRI P J. Macroalgal fouling communities as indicators of environmental change: Potential applications for water quality monitoring. Journal of the Marine Biological Association of the United Kingdom, 2018, 98(7): 1581-1588 DOI:10.1017/S0025315417001102 |
FUNAMOTO T, AOKI I, WADA Y. Reproductive characteristics of Japanese anchovy, Engraulis japonicus, in two bays of Japan. Fisheries Research, 2004, 70(1): 71-81 DOI:10.1016/j.fishres.2004.06.017 |
GARRISON L P. Spatial patterns in species composition in the northeast United States continental shelf fish community during 1966-1999. Spatial Processes and Management of Marine Populations, 2001, 513-559 |
GOWER J C. A general coefficient of similarity and some of its properties. Biometrics, 1971, 27: 857-871 DOI:10.2307/2528823 |
HOOPER D U, DUKES J S. Overyielding among plant functional groups in a long-term experiment. Ecology Letters, 2004, 7(2): 95-105 DOI:10.1046/j.1461-0248.2003.00555.x |
HOVENKAMP F. Growth-dependent mortality of larval plaice Pleuronectes platessa in the North Sea. Marine Ecology Progress Series, 1992, 82: 95-101 DOI:10.3354/meps082095 |
LEI J L, FAN N C, ZHENG C W. A preliminary observation on the morphological characteristics of embryos, larvae and juveniles of Nibea albiflora (Richardson). Marine Fisheries Research, 1981(1): 77-84 [雷霁霖, 樊宁臣, 郑澄伟. 黄姑鱼(Nibea albiflora Richardson)胚胎及仔、稚鱼形态特征的初步观察. 海洋水产研究, 1981(1): 77-84] |
LI J D, ZHAO Q, LIU J J, et al. Study on the characteristics of fishery biological community structure and factors influencing biomass increments in different artificial reefs in the Yellow Sea and Bohai Sea. Journal of Fishery Sciences of China, 2023, 30(3): 371-383 [李建都, 赵祺, 刘晋冀, 等. 黄渤海不同人工鱼礁区渔业生物群落结构特征及生物增量影响要素. 中国水产科学, 2023, 30(3): 371-383] |
LUAN Q S, KANG Y D, WANG J. Long-term changes in the phytoplankton community in the Bohai Sea (1959~2015). Progress in Fishery Sciences, 2018, 39(4): 9-18 [栾青杉, 康元德, 王俊. 渤海浮游植物群落的长期变化(1959~2015). 渔业科学进展, 2018, 39(4): 9-18] |
MAYFIELD M M, BONI M F, DAILY G C, et al. Species and functional diversity of native and human-dominated plant communities. Ecology, 2005, 86(9): 2365-2372 |
MOUILLOT D, DUMAY O, TOMASINI J A. Limiting similarity, niche filtering and functional diversity in coastal lagoon fish communities. Estuarine Coastal and Shelf Science, 2007, 71(3/4): 443-456 |
NIU M, JIN X, LI X, et al. Effects of spatio-temporal and environmental factors on distribution and abundance of wintering anchovy Engraulis japonicus in central and southern Yellow Sea. Chinese Journal of Oceanology and Limnology, 2014, 32(3): 565-575 |
OKIYAMA M. An atlas of the early stage fishes in Japan. second edition. Tokyo: Tokai University Press, 2014 [冲山宗雄. 日本产稚鱼图鉴. 2版. 东京: 东海大学出版会, 2014]
|
PAN L, HU F W, GAO F X, et al. Study of artificial breeding and seedling culture technique for greenling Hexagrammos otakii. Marine Sciences, 2012, 36(12): 39-44 [潘雷, 胡发文, 高凤祥, 等. 大泷六线鱼人工繁殖及育苗技术初步研究. 海洋科学, 2012, 36(12): 39-44] |
PINHEIRO H T, GOODBODY-GRINGLEY G, JESSUP M E, et al. Upper and lower mesophotic coral reef fish communities evaluated by underwater visual censuses in two Caribbean locations. Coral Reefs, 2016, 35: 139-151 |
PINKAS L. Food habits of albacore, bluefin tuna and bonito in California waters. California Department of Fish and Game, Fish Bulletin, 1971, 152: 1-105 |
RUI Y, JIANG R J, WANG H X, et al. Characteristics of fish community structure and its relationship with environmental factors in Dachenyang spawning ground reserve. Journal of Fisheries of China, 2022, 46(6): 995-1007 [芮银, 蒋日进, 王好学, 等. 大陈洋产卵场保护区鱼类群落结构特征及其与环境因子的关系. 水产学报, 2022, 46(6): 995-1007] |
SABATÉS A, OLIVAR M P, SALAT J, et al. Physical and biological processes controlling the distribution of fish larvae in the NW Mediterranean. Progress in Oceanography, 2007, 74(2/3): 355-376 |
SUN X, ZHANG Y L, XU B D, et al. Habitat suitability of Cynoglossus joyneri spawning grounds in Haizhou Bay and adjacent waters. Journal of Fishery Sciences of China, 2020, 27(12): 1505-1514 [孙霄, 张云雷, 徐宾铎, 等. 海州湾及邻近海域短吻红舌鳎产卵场的生境适宜性. 中国水产科学, 2020, 27(12): 1505-1514] |
TAKASUKA A, OOZEKI Y, AOKI I. Optimal growth temperature hypothesis: Why do anchovy flourish and sardine collapse or vice versa under the same ocean regime. Canadian Journal of Fisheries and Aquatic Sciences, 2007, 64(5): 768-776 |
TANG Q S. On the carrying capacity and its study. Marine Fisheries Research, 1996, 17(2): 1-6 [唐启升. 关于容纳量及其研究. 海洋水产研究, 1996, 17(2): 1-6] |
TARIMO B, WINDER M, MTOLERA M S P, et al. Seasonal distribution of fish larvae in mangrove-seagrass seascapes of Zanzibar (Tanzania). Scientific Reports, 2022, 12(1): 4196 |
VILLÉGER S, MASON N W H, MOUILLOT D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301 |
WAN R J, JIANG Y W. Studies on the ecology of eggs and larvae of Osteichthyes in the Yellow Sea. Marine Fisheries Research, 1998b, 19(1): 60-73 [万瑞景, 姜言伟. 黄海硬骨鱼类鱼卵、仔稚鱼及其生态调查研究. 海洋水产研究, 1998b, 19(1): 60-73] |
WAN R J, JIANG Y W. The distribution and variation of eggs and larvae of Osteichthyes in the Bohai Sea. Journal of Fishery Sciences of China, 1998a, 5(1): 44-51 [万瑞景, 姜言伟. 渤海硬骨鱼类鱼卵和仔稚鱼分布及其动态变化. 中国水产科学, 1998a, 5(1): 44-51] |
WAN R J, ZHANG R Z. Fish eggs, larvae and juveniles in the offshore and adjacent waters of China. Shanghai: Shanghai Scientific and Technical Publishers, 2016 [万瑞景, 张仁斋. 中国近海及其邻近水域鱼卵与仔稚鱼. 上海: 上海科学技术出版社, 2016]
|
WAN R, ZHANG T Z, LI Z G, et al. Spatial distribution and inter-annual variability of spawning grounds of Cynoglossus joyneri in the Yellow Sea coastal waters in summer. Chinese Journal of Applied Ecology, 2020, 31(3): 1023-1032 [万荣, 张同征, 李增光, 等. 黄海近岸海域短吻红舌鳎夏季产卵场的空间分布及其年际变化. 应用生态学报, 2020, 31(3): 1023-1032] |
WATSON D F, PHILIP G M. A refinement of inverse distance weighted interpolation. Geo-Processing, 1985, 2: 315-327 |
YASUDA T, KINOSHITA J, NIINO Y, et al. Vertical migration patterns linked to body and environmental temperatures in chub mackerel. Progress in Oceanography, 2023, 213: 103017 |
YU H, YU H, ITO S, et al. Potential environmental drivers of Japanese anchovy (Engraulis japonicus) recruitment in the Yellow Sea. Journal of Marine Systems, 2020, 212: 103431 |
YU X G, DONG J, LI Y P, et al. Species composition and protection of fish eggs and larvae in the coastal waters in Liaodong Bay. Journal of Dalian Ocean University, 2018, 33(3): 370-378 [于旭光, 董婧, 李轶平, 等. 辽东湾近海鱼卵、仔稚鱼种类组成和保护分析. 大连海洋大学学报, 2018, 33(3): 370-378] |
YU X G, LI Y P, YAN J Y, et al. Species composition and quantity distribution of fish eggs and larvae in coastal waters of the north Yellow Sea. Marine Fisheries, 2020, 42(6): 659-671 [于旭光, 李轶平, 燕金宜, 等. 黄海北部近岸水域鱼卵、仔稚鱼种类组成与数量分布. 海洋渔业, 2020, 42(6): 659-671] |
ZHANG R L, LIU H, SUN D Y, et al. Characteristics of benthic fishery community at natural reefs and artificial reefs located in Yantai offshore area. Oceanologia et Limnologia Sinica, 2021, 52(3): 697-707 [张荣良, 刘辉, 孙东洋, 等. 烟台近岸人工鱼礁与自然岩礁底层渔业生物群落特征对比分析. 海洋与湖沼, 2021, 52(3): 697-707] |
ZHANG R Z, LU S F, ZHAO C Y, et al. Fish eggs and larvae in the coastal waters of China. Shanghai: Shanghai Scientific and Technical Publishers, 1985 [张仁斋, 陆穗芬, 赵传絪, 等. 中国近海鱼卵与仔鱼. 上海: 上海科学技术出版社, 1985]
|
ZHANG X Z, WANG J, XU B D, et al. Spatio-temporal variations of functional diversity of fish communities in Haizhou Bay. Chinese Journal of Applied Ecology, 2019, 30(9): 3233-3244 [张晓妆, 王晶, 徐宾铎, 等. 海州湾鱼类群落功能多样性的时空变化. 应用生态学报, 2019, 30(9): 3233-3244] |
ZHANG Y X, BIAN X D, SHAN X J, et al. Spatio-temporal distribution of chub mackerel (Scomber japonicus) spawning grounds and its relationship with environmental factors in the offshore waters of Yantai-Weihai. Journal of Fishery Sciences of China, 2022a, 29(4): 618-632 [张雨轩, 卞晓东, 单秀娟, 等. 烟威近岸水域鲐产卵场时空分布及其与环境因子的关系. 中国水产科学, 2022a, 29(4): 618-632] |
ZHANG Y X, BIAN X D, SHAN X J, et al. Community structure and suitable habitat of the early life stages of marine fish in the Yantai-Weihai offshore waters. Progress in Fishery Sciences, 2022b, 43(6): 148-167 [张雨轩, 卞晓东, 单秀娟, 等. 烟威近岸水域鱼类早期资源群落结构及适宜产卵生境. 渔业科学进展, 2022b, 43(6): 148-167] |
ZHAO C Y. Marine fishery resources of China. Hangzhou: Zhejiang Science and Technology Publishing House, 1990: 34-109 [赵传 . 中国海洋渔业资源. 杭州: 浙江科学技术出版社, 1990: 34-109]
|
ZHAO Y S, SHAN X J, YANG T, et al. Biodiversity characteristics of bottom fishery communities in the sea adjacent to the Miaodao Archipelago. Progress in Fishery Sciences, 2022, 43(6): 132-147 [赵永松, 单秀娟, 杨涛, 等. 庙岛群岛毗邻海域底层渔业生物群落多样性特征. 渔业科学进展, 2022, 43(6): 132-147] |
ZOU J Y, LIU S D, ZHANG C L, et al. Seasonal and spatial changes in functional diversity of fish communities in the adjacent waters of the Changshan Islands. Haiyang Xuebao, 2023, 45(1): 13-24 [邹建宇, 刘淑德, 张崇良, 等. 长山列岛邻近海域鱼类群落功能多样性的季节和空间变化. 海洋学报, 2023, 45(1): 13-24] |