



海洋牧场是改善海域生态环境,养护渔业资源,实现渔业资源可持续利用的渔业模式,目前在国内外被广泛应用(陈丕茂等, 2019; 杨红生等, 2019)。根据山东省地方标准《海洋牧场建设规范第1部分:术语和分类》(DB37/T 2982.1-2017)可知,山东的海洋牧场基于生产方式分为投礁型、游钓型、底播型、田园型和装备型海洋牧场,其中,田园型海洋牧场是指以筏式养殖设施等立体生态方式为主的海洋牧场。
筏式养殖设施是我国近岸海域牡蛎、扇贝和贻贝等双壳类筏式养殖所需要构建的养殖框架(丁刚等, 2013)。在双壳类养殖过程中,筏式养殖设施为海洋中其他生物提供了良好的附着基质,出现大量附着生物在养殖设施上栖息和生存的现象(水柏年等, 2008)。相关研究表明,在牡蛎、扇贝和贻贝等贝类筏式养殖区,海鞘、藻类等附着生物的存在会导致养殖吊笼网孔堵塞,使养殖贝类因为缺氧而死亡(张俊波等, 2021),海绵类附着动物的活动会软化筏式养殖设施,缩短筏式养殖设施的使用寿命(Fitridge et al, 2012; 刘义等, 2022),从而对贝类筏式养殖产生不利影响,造成经济损失。然而,Leblanc等(2003)对加拿大贻贝养殖区的附着生物进行调查发现,该贻贝养殖区释放的无机氮有60%来自于附着生物的排泄。Mazouni等(2001)对法国牡蛎养殖区的附着生物调查发现,悬浮生物(指牡蛎以及养殖筏架上附着生物的统称)与养殖水体之间的营养盐交换速率主要受附着生物的影响。这说明,附着生物的存在会加快氮磷等营养盐循环。相关研究表明,无机氮、无机磷等营养盐的快速循环,促进了浮游植物的生长,提高了初级生产力(齐占会等, 2021),为牡蛎、扇贝和贻贝等双壳类提供更多的饵料,增加双壳类的肥满度(Lacoste et al, 2015),对筏式养殖区生态环境是有利的。不同筏式养殖区域的附着生物种类数和生物量及其与环境因子的关系存在特异性,因此,有必要针对筏式养殖区展开附着生物基础研究,为双壳类的健康养殖提供参考依据,进而为探究筏式养殖设施上附着生物对生态环境的影响提供基础资料。
综上所述,本研究基于乳山田园型海洋牧场牡蛎筏式养殖设施上4个季度的附着生物调查数据,探究筏式养殖设施上附着生物的季节变化及其与环境因子的关系。
1 材料与方法 1.1 调查海域和内容调查海域为乳山华信田园型海洋牧场区(图 1),该海洋牧场属于省级海洋牧场,位于山东省乳山市海洋所镇波浪滩码头以南6 n mile处。该海域为开放海域,受陆地影响较小,平均水深为21 m。牡蛎养殖面积为297 hm2,总共布设养殖筏架1 200个,每个养殖筏架呈东西方向排列,2个筏架之间的距离为20 m,每个筏架长约70 m,挂设牡蛎养殖吊笼50~60笼,每个吊笼长1.5 m,直径为35 cm,养殖水深为水下4~5 m处。

|
图 1 调查站位 Fig.1 Investigation sites |
调查时间为2022年7月(夏季)、10月(秋季)、2023年1月(冬季)和4月(春季)。每次调查在牡蛎养殖区共设置4个固定站位。牡蛎单个养殖筏架由木桩、撅绳、浮子、吊笼、吊绳和浮绠六部分组成(图 2)。

|
图 2 单个牡蛎养殖筏架结构示意图 Fig.2 Structure diagram of a single oyster culture raft |
每个站位进行附着生物调查时,分别在浮子、浮绠、吊笼3个不同部位进行采样,采样面积分别为10 cm×10 cm、283 cm2(长度30 cm,直径3 cm处)、20 cm ×20 cm,对采集的样品进行装瓶编号,再用5%甲醛溶液固定保存,带回实验室进行种类鉴定与计数。
现场测定温度、盐度、溶解氧、酸碱度、透明度等,实验室测定样品的化学需氧量、叶绿素、硅酸盐、磷酸盐、无机氮等。样品的采集与处理根据《海洋监测规范》(GB 17378.4-2007)和《海洋调查规范》(GB/T 12763.6-2007)进行。
1.2 数据处理与分析使用Margalef丰富度指数(D) (Margalef, 1957)、Shannon-Wiener多样性指数(H') (Shannon et al, 1964)和Pielou均匀度指数(J') (Levins, 1970)分析附着生物群落特征。
采用相对重要性指数(IRI)计算附着生物的优势种。
| $ {\text{IRI}} = (N + W) \times F \times {10^4} $ | (1) |
式中,N代表某一物种的丰度占总丰度的百分比;W代表某一物种的生物量占总生物量的百分比;F为某一物种的出现频率;本研究将IRI≥1 000定义为优势种。
附着生物量计算公式:
| $ P = W/S $ | (2) |
式中,P为附着生物量(g/m2),W为附着生物取样重量(g),S为取样面积(m2)。
使用Primer 6.0软件对附着生物进行聚类分析和Simper相似性百分比分析。
利用R语言程序包进行Permanova分析,检验不同站位、季节之间的环境因子、生物群落的差异显著性;进行非度量多维尺度分析(NMDS分析);进行Spearman分析和去趋势对应分析(DCA),再根据分析结果的最长轴选择典范对应分析(CCA)或者冗余分析(RDA),分析环境因子与生物群落优势种之间的关系;进行蒙特卡洛置换检验,检验各环境因子对各生物群落影响的显著性。本研究的显著性水平为P < 0.05,极显著性水平为P < 0.01。
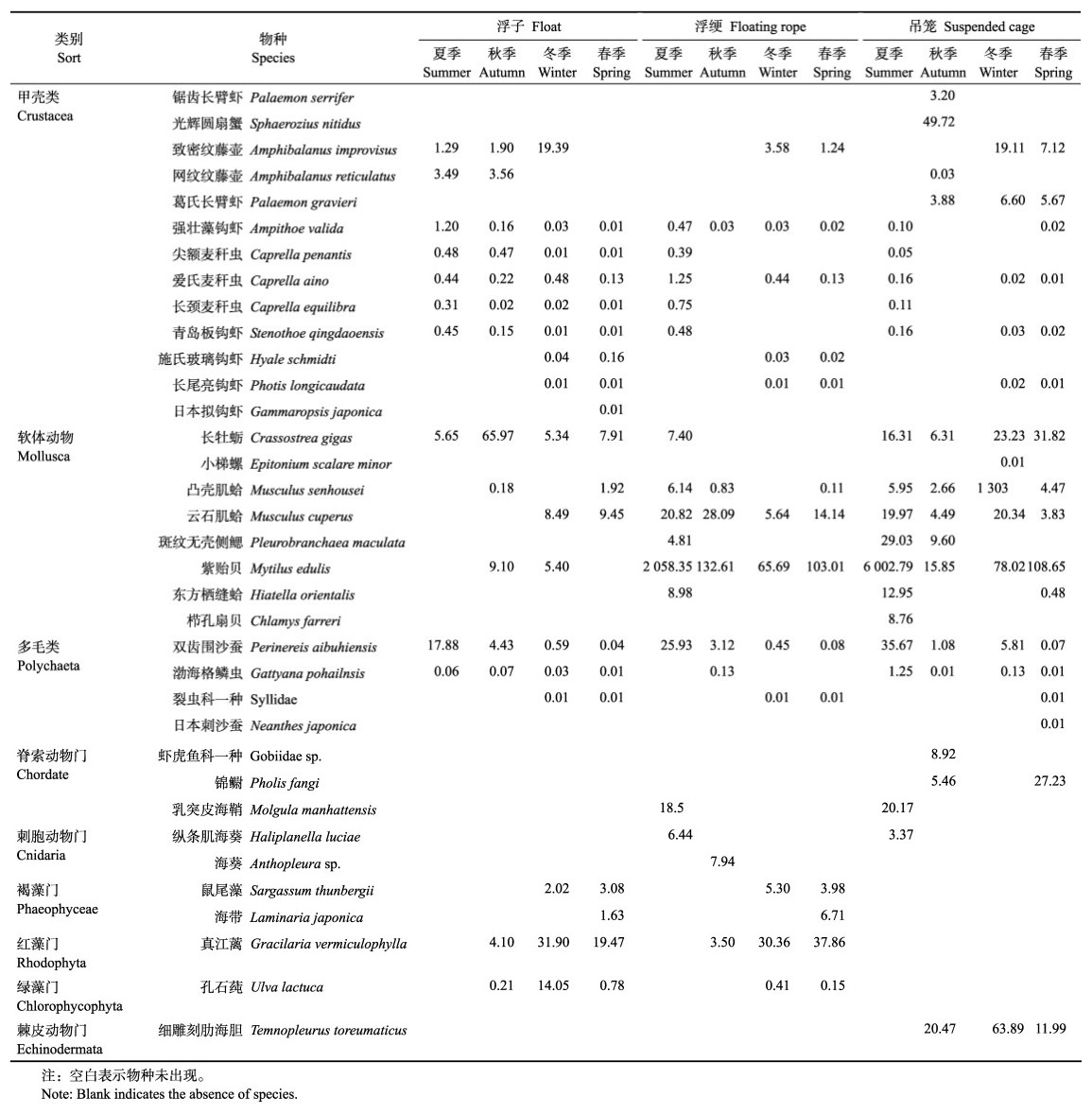
2 结果 2.1 附着生物的种类组成和丰度变化共发现附着生物8大类35种,其中,甲壳类(Crustacea) 13种、软体动物(Mollusca) 8种、多毛类(Polychaeta) 4种、脊索动物(Chordate) 3种、刺胞动物(Cnidaria) 2种、褐藻门(Phaeophyceae) 2种、红藻门(Rhodophyta) 1种、绿藻门(Chlorophycophyta) 1种、棘皮动物(Echinodermata) 1种,见表 1。附着生物的种类数在季节和3个采样部位之间均存在显著差异(P < 0.05, Permanova),在4个调查站位之间无显著差异(P > 0.05, Permanova)。浮绠上附着生物种类数在夏、春季最多,各为14种,其次为冬季12种和秋季8种;吊笼上附着生物种类数从高到低依次为春季、夏季、秋季、冬季;浮子上附着生物种类数从高到低依次为春季、冬季、秋季和夏季(图 3a)。
|
|
表 1 附着生物种类组成及生物量/(g/m2) Tab.1 The species composition and biomass of fouling organisms/(g/m2) |

|
图 3 附着生物的种类数(a)和生物量(b) Fig.3 The species number (a) and biomass (b) of fouling organisms |
从生物量变化情况来看(图 3b),夏季附着生物生物量极显著高于其他季节(P < 0.01,Permanova),除夏季外,相同季节内3个不同采样部位的附着生物量无显著差异(P > 0.05,Permanova)。不同采样部位中吊笼上的附着生物量变化范围最大,其次为浮绠和浮子,其生物量变化范围分别为31.48~1 645.82、23.22~ 580.76和5.76~30.12 g/m2。
2.2 附着生物的群落结构季节变化以及多样性分析聚类分析和NMDS分析结果表明,相同采样部位的附着生物可以在较高水平上聚为一类,而不同季节之间的附着生物没有明显的规律。夏季浮绠、吊笼和浮子之间的附着生物群落结构相似度最高,相似度可达到80%以上,冬春两季相同采样部位的附着生物群落结构相似度较高,秋季3个不同采样部位的相似度最低(图 4)。Simper相似性百分比分析结果显示,附着生物种类组成在不同采样部位之间的相似度较低,在不同季节之间冬季和春季的相似度为80.17%,其余季节之间相似度都在50%左右(表 2)。

|
图 4 附着生物的聚类分析和NMDS分析 Fig.4 The cluster analysis and NMDS analysis of fouling organisms SC:吊笼;FR:浮绠;FL:浮子;SU:夏季;AU:秋季;WI:冬季;SP:春季。下同。 SC: Suspended cage; FR: Floating rope; FL: Float; SU: Summer; AU: Autumn; WI: Winter; SP: Spring. The same below. |
|
|
表 2 造成不同季节之间和不同采样部位之间附着生物种类差异的累加贡献率前30%的物种 Tab.2 Species in the top 30% of the cumulative contribution to the differences in fouling organisms between different seasons and different sampling sites |
4个季度调查期间附着生物的丰富度指数、均匀度指数和多样性指数的变化范围分别为1.38~3.61、0.33~0.92和0.88~2.58,丰富度指数最低值出现在夏季浮子上,均匀度指数和多样性指数在夏季吊笼上最低,3个指数均在春季吊笼上达到最高值(图 5)。Permanova分析结果显示,3个指数在不同季节和不同采样部位之间均存在显著差异(P < 0.05)。

|
图 5 附着生物的群落参数 Fig.5 Community parameters of fouling organisms |
调查期间共发现附着生物优势种21种(表 3),其中浮绠和吊笼上各出现11种优势种,浮子上出现12种优势种,优势种组成在不同采样部位存在明显差异,且存在一定的季节变化。紫贻贝为浮绠和吊笼上的常年优势种,夏季浮子上的主要优势种为双齿围沙蚕(Perinereis aibuhiensis),秋季浮子上的主要优势种为长牡蛎(Crassostrea gigas),冬季和春季浮子上的主要优势种为真江蓠(Gracilaria vermiculophylla),爱氏麦秆虫(Caprella aino)为浮子上的常年优势种。
|
|
表 3 不同季节和不同采样部位之间附着生物优势种编号及其相对重要性指数 Tab.3 The code and relative importance index of the fouling organisms dominant species between different seasons and different sampling stations |
4个季度调查期间的海域环境因子中(表 4),温度和透明度存在显著季节差异,变化规律相同,由高到低均为夏季、秋季、春季和冬季,溶解氧(DO)的季节变化规律由高到低依次为冬季、春季、夏季和秋季;盐度和pH在4个季度调查期间基本不变;叶绿素、化学需氧量(COD)和营养盐在季节间没有明显的变化规律。
|
|
表 4 调查海域环境因子时空分布 Tab.4 Temporal and spatial characteristics of environment variables |
将环境因子与附着生物优势种进行冗余分析(RDA)、Spearman相关性分析和蒙特卡洛置换检验。结果显示(图 6),冗余分析前2个轴的解释量分别为98.86%和0.96%,环境因子对附着生物优势种的总解释量为69.92%,影响显著性从高到低依次为透明度、温度、硅酸盐、叶绿素、DO、COD、磷酸盐和pH,其中,盐度和无机氮对附着生物优势种无显著性影响。紫贻贝(F9)作为调查海域的第一优势种,其与透明度和温度呈正相关,与叶绿素和DO呈负相关。冬季和春季的主要优势种真江蓠(F3)与透明度和温度呈负相关,与叶绿素和DO呈正相关。不同物种与环境因子之间的相关性也不相同。

|
图 6 附着生物与环境因子的冗余分析以及相关性分析 Fig.6 Redundancy analysis and correlation analysis of fouling organisms with environmental factors F1~F21:附着生物优势种编号。 F1~F21: The code of fouling organisms' dominant species. |
田园型海洋牧场牡蛎筏式养殖设施上共调查发现了附着生物35种,其中,甲壳类种类数最多,其次为软体动物。从空间尺度来看,浮子、浮绠和吊笼3个不同采样部位之间的附着生物优势种类和生物量也互不相同,没有明显的规律变化,紫贻贝为该调查海域筏式养殖设施上的绝对优势物种。严涛等(2008)对黄渤海污损生物进行综合分析表明,在离岸较远的开阔海域,紫贻贝将在附着生物中占据优势地位。本研究结果与其一致,紫贻贝为浮绠和吊笼上的常年优势种,其没有在浮子上成为优势种可能与水深和附着基材质太光滑有关(刘明坤等, 2022)。从时间尺度来看,春季附着生物种类数最多,秋季种类数最少,甲壳类、软体动物及多毛类在各个季节均有分布。造成秋季附着生物种类数最少的原因可能是因为从每年9月开始,乳山的养殖户会从荣成等地区收购成熟期的牡蛎苗,在乳山海域进行育肥工作,在此期间,渔民会悬挂新的养殖吊笼来养殖牡蛎,因此,秋季是所有海洋生物附着的初始阶段,所以该季节附着生物种类数最少,还没有形成稳定的生物群落。
从丰度和生物量上来看,在夏季,紫贻贝丰度和生物量均高于其他生物,说明该季节紫贻贝大量繁殖且个体较大,而在秋冬春三季,由于温度降低和人工清理等因素的共同作用,紫贻贝丰度和生物量迅速下降,这就造成了夏季浮绠和吊笼上的附着生物量显著高于其他季节。同时,由于紫贻贝大量繁殖,占据了绝大部分附着基,导致其他附着生物无法生存,因此,夏季多样性指数和均匀度指数均低于其他季节。该结果与齐占会等(2010、2011)对桑沟湾贝藻养殖区以及青岛流清河湾扇贝养殖区的附着生物调查结果相一致,其挂网研究发现,在夏季,紫贻贝为附着生物群落的绝对优势种,而且多样性较低。因为紫贻贝属于暖水性物种(孙大鹏等, 2010),所以海水温度也是影响其附着生物量的重要环境因素,当温度较低时,紫贻贝优势种地位降低,物种多样性增加。
孔石莼(Ulva lactuca)、真江蓠等海藻在冬春两季成为浮子和浮绠上的优势种,可能是这些海藻更喜欢低温环境(Fan et al, 2015)。研究表明,大型海藻的生长和光合作用需要吸收无机氮、磷酸盐和硅酸盐等无机物(章守宇等, 2019),所以导致冬季和春季的营养盐浓度较低。同时,大型海藻作为饵料可以促进腹足类和甲壳类生物的生长(郑新庆等, 2013; 薛素燕等, 2022),增加了冬春两季的物种多样性。除了直接被植食性动物摄食之外,大型海藻产生的碎屑也是其他海洋生物的食物来源之一(吴程宏等, 2017),而且藻类碎屑的增加也会导致海水透明度降低,这可能是调查期间冬春两季透明度较低的原因。相较于章守宇等(2021)对枸杞岛贻贝养殖区的大型附着海藻生物量的调查,本次调查海域大型藻类生物量较低,可能是因为该海域处于开放海域,且牡蛎养殖设施的构建相对于枸杞岛近岸贻贝养殖设施较为松散(枸杞岛贻贝养殖区布设近2 000个100 m×50 m的养殖单元,每个养殖单元由将近400个浮筏、2 000条贻贝苗绳以及大量撅绳和浮绠组成),对水流交换的阻滞作用不明显,使得牡蛎养殖区内的藻类孢子不容易附着在养殖设施上,所以大型藻类生物量较低。
除温度外,海水盐度对附着生物的分布也起着决定性作用(王晓波等, 2012),但在本研究中,冗余分析结果表明盐度对附着生物没有显著性影响,可能是因为该调查海域海水盐度在4个季度调查期间基本不变,一直处于高盐度状态,所以该海域附着生物都适宜在高盐度海水中生存,不会因为海水盐度的轻微变化而改变。陈义华等(2019)对海州湾延绳式牡蛎养殖区附着生物调查发现,高盐度的海水适合海鞘类生物的生存。陈凯等(2023)研究表明,海鞘生物的空间分布存在明显的地域特征,大多数海鞘更适合在热带海域生存。因此,在本研究中未发现大量海鞘类生物的存在,这可能是由于调查海域的海水温度较低所导致的。
将本次筏式养殖设施上附着生物的调查结果与詹启鹏等(2022)对人工鱼礁的附着生物群落结构调查对比可知,人工鱼礁上的附着生物以节肢动物门为主,优势种为脉红螺(Rapana venosa)以及虾蟹类等大型物种,而筏式养殖设施的附着生物以麦秆虫和钩虾等小型物种为主,主要原因可能是人工鱼礁受人为因素影响较小,且其附着基较大,可供大型物种栖息繁殖,而筏式养殖设施受人为因素影响较大,渔民会定期对养殖筏架和养殖吊笼进行清理,且附着基较小,所以不会出现大型物种。
根据本研究所调查的附着生物季节变化规律,建议渔民在春夏两季适当加快养殖吊笼更换频率,可最大限度地保证牡蛎的存活率,避免牡蛎大量死亡,造成经济损失,更换下来的网衣可通过暴晒法和机械敲打清除网衣上附着的紫贻贝等软体动物。除此之外,在养殖过程中,也可以使用环保无毒型防污涂料来浸泡养殖吊笼,减少生物附着,从而降低附着生物对牡蛎养殖的危害。然而,虽然如今对新型环保型防污涂料的研究有很多,但缺少环保型防污涂料的测试实验,无法保障环保型防污涂料的实际应用效果。因此,亟需科研人员对新型防污涂料开展防生物附着测试实验,以期寻找无毒、有效、价格适宜的新型环保型涂料,保障渔民的经济利益。
CHEN K, WANG S T, YUAN K X. Distribution characteristics and prevention of fouling organisms in coastal areas of China. Journal of Aquaculture, 2023(11): 40–45, 62 [陈凯, 王胜通, 元轲新. 我国沿海污损生物分布特点及防治措施. 水产养殖, 2023(11): 40–45, 62] |
CHEN P M, SHU L M, YUAN H R, et al. Review on development, definition and classification of marine ranching in domestic and overseas. Journal of Fisheries of China, 2019, 43(9): 1851-1869 [陈丕茂, 舒黎明, 袁华荣, 等. 国内外海洋牧场发展历程与定义分类概述. 水产学报, 2019, 43(9): 1851-1869] |
CHEN Y H, DONG Z G, LI X Y, et al. Study on fouling organisms in Pacific oysters (Crassostrea gigas) culture area of the Haizhou Bay. Genomics and Applied Biology, 2019, 38(4): 1572-1579 [陈义华, 董志国, 李晓英, 等. 海州湾长牡蛎养殖区污损生物研究. 基因组学与应用生物学, 2019, 38(4): 1572-1579] |
DING G, WU H Y, GUO P P, et al. Evolution and development trend of marine raft cultivation model in China. Chinese Fisheries Economics, 2013, 31(1): 164-169 [丁刚, 吴海一, 郭萍萍, 等. 我国海上筏式养殖模式的演变与发展趋势. 中国渔业经济, 2013, 31(1): 164-169] |
FAN S L, FU M Z, WANG Z L, et al. Temporal variation of green macroalgal assemblage on Porphyra aquaculture rafts in the Subei Shoal, China. Estuarine, Coastal and Shelf Science, 2015, 163: 23-28 DOI:10.1016/j.ecss.2015.03.016 |
FITRIDGE I, DEMPSTER T, GUENTHER J, et al. The impact and control of biofouling in marine aquaculture: A review. Biofouling, 2012, 28(7): 649-669 DOI:10.1080/08927014.2012.700478 |
LACOSTE E, GAERTNER-MAZOUNI N. Biofouling impact on production and ecosystem functioning: A review for bivalve aquaculture. Reviews in Aquaculture, 2015, 7(3): 187-196 DOI:10.1111/raq.12063 |
LEBLANC A R, LANDRY T, MIRON G. Fouling organisms of the blue mussel Mytilus edulis: Their effect on nutrient uptake and release. Journal of Shellfish Research, 2003, 22(3): 633-638 |
LEVINS R. An introduction to mathematical ecology. Evolution, 1970, 24(2): 482 |
LIU M K, QUE H Y, ZHANG G F, et al. The current standing of the Mediterranean mussel industry, the obstacles posed and its potential restoration. Marine Sciences, 2022, 46(3): 135-144 [刘明坤, 阙华勇, 张国范, 等. 紫贻贝养殖产业的现状、问题与对策. 海洋科学, 2022, 46(3): 135-144] |
LIU Y, LIANG Z R, YU W W, et al. Effects of cultivation area of different offshore distances on the agronomic traits and breeding rope aging of Saccharina japonica. Progress in Fishery Sciences, 2022, 43(3): 156-164 [刘义, 梁洲瑞, 余雯雯, 等. 不同离岸距离的养殖海区对海带养殖性状及苗绳老化的影响. 渔业科学进展, 2022, 43(3): 156-164] |
MARGALEF R. Information theory in ecology. General Systems, 19587
|
MAZOUNI N, GAERTNER J C, DESLOUS-PAOLI J M. Composition of biofouling communities on suspended oyster cultures: An in situ study of their interactions with the water column. Marine Ecology Progress Series, 2001, 214(1): 93-102 |
QI Z H, FANG J G, ZHANG J H, et al. Seasonal succession of fouling communities in the poly-culture area of scallop Chlamys farreri and kelp Laminaria japonica in Sanggou Bay. Progress in Fishery Sciences, 2010, 31(4): 72-77 [齐占会, 方建光, 张继红, 等. 桑沟湾贝藻养殖区附着生物群落季节演替研究. 渔业科学进展, 2010, 31(4): 72-77] |
QI Z H, SHI R J, YU Z H, et al. Review of influences of filter-feeding bivalves aquaculture on planktonic community. South China Fisheries Science, 2021, 17(3): 115-121 [齐占会, 史荣君, 于宗赫, 等. 滤食性贝类养殖对浮游生物的影响研究进展. 南方水产科学, 2021, 17(3): 115-121] |
QI Z H, WANG J, FANG J G, et al. The seasonal succession of fouling organisms in Liuqing River estuary. Progress in Fishery Sciences, 2011, 32(4): 86-92 [齐占会, 王珺, 方建光, 等. 流清河湾扇贝养殖区夏季附着生物特征研究. 渔业科学进展, 2011, 32(4): 86-92] |
SHANNON C E, WEAVER W. The mathematical theory of communication. Urbana: University of Illinois Press, 1964
|
SHUI B N, GUO D F. Seasonal variation analysis of fouling organisms on offshore cages in Zhoushan. South China Fisheries Science, 2008, 4(4): 36-41 [水柏年, 郭迪飞. 舟山深水网箱污损生物季节性变化分析. 南方水产, 2008, 4(4): 36-41] |
SUN D P, SHAO M Y, MA Y B, et al. Research on species composition and distribution of large-scale fouling organisms on scallop farming cages using test panels. Periodical of Ocean University of China (Natural Science), 2010, 40(9): 84-90 [孙大鹏, 邵明瑜, 马玉彬, 等. 扇贝养殖笼大型污损生物种类组成及分布的挂板研究. 中国海洋大学学报(自然科学版), 2010, 40(9): 84-90] |
WANG X B, LONG S Q, HE D H, et al. Distribution characteristics of fouling organisms and their relationship with environmental facrors in Zhujiajian waters, Zhoushan. Journal of Marine Sciences, 2012, 30(1): 41-50 [王晓波, 龙绍桥, 何东海, 等. 舟山朱家尖海域污损生物的分布特征及其与环境因子的关系. 海洋学研究, 2012, 30(1): 41-50] |
WU C H, ZHANG S Y, ZHOU X J, et al. Identifying sources of sedimentary organic matter in the rocky reef seaweed bed. Journal of Fisheries of China, 2017, 41(8): 1246-1255 [吴程宏, 章守宇, 周曦杰, 等. 岛礁海藻场沉积有机物来源辨析. 水产学报, 2017, 41(8): 1246-1255] |
XUE S Y, MAO Y Z, LI J Q, et al. The feeding selectivty of amphipod Eogammarus possjeticus on macroalgae and its potential carbon sink analysis. Progress in Fishery Sciences, 2022, 43(5): 40-48 [薛素燕, 毛玉泽, 李加琦, 等. 钩虾对大型海藻的摄食选择及其潜在的碳汇分析. 渔业科学进展, 2022, 43(5): 40-48] |
YAN T, CAO W H. Ecology of marine fouling organism in Huanghai and Bohai Seas. Journal of Marine Sciences, 2008, 26(3): 107-118 [严涛, 曹文浩. 黄、渤海污损生物生态特点及研究展望. 海洋学研究, 2008, 26(3): 107-118] |
YANG H S, ZHANG S Y, ZHANG X M, et al. Strategic thinking on the construction of modern marine ranching in China. Journal of Fisheries of China, 2019, 43(4): 1255-1262 [杨红生, 章守宇, 张秀梅, 等. 中国现代化海洋牧场建设的战略思考. 水产学报, 2019, 43(4): 1255-1262] |
ZHAN Q P, SUN X, DONG J Y, et al. Community structure and functional traits of attached organisms on an artificial reef near Furong Island, Laizhou Bay. Journal of Fishery Sciences of China, 2022, 29(5): 704-719 [詹启鹏, 孙昕, 董建宇, 等. 莱州芙蓉岛海域人工鱼礁附着生物的群落结构和功能性状. 中国水产科学, 2022, 29(5): 704-719] |
ZHANG J B, SUN M S, WAN R. Research progress of biofouling prevention techniques for marine fishery equipment. Journal of Fishery Sciences of China, 2021, 28(11): 1489-1503 [张俊波, 孙名山, 万荣. 海洋渔业装备生物附着控制技术研究进展. 中国水产科学, 2021, 28(11): 1489-1503] |
ZHANG S Y, CUI X, WANG Z H, et al. Community structure of epiphytic macroalgae on mussel culture rafts in Gouqi Island. Journal of Fisheries of China, 2021, 45(5): 726-739 [章守宇, 崔潇, 汪振华, 等. 枸杞岛贻贝养殖筏架附着海藻的群落结构. 水产学报, 2021, 45(5): 726-739] |
ZHANG S Y, LIU S R, ZHOU X J, et al. Ecological function of seaweed-formed habitat and discussion of its application to sea ranching. Journal of Fisheries of China, 2019, 43(9): 2004-2014 [章守宇, 刘书荣, 周曦杰, 等. 大型海藻生境的生态功能及其在海洋牧场应用中的探讨. 水产学报, 2019, 43(9): 2004-2014] |
ZHENG X Q, HUANG L F, LI Y C, et al. The feeding selectivity of an herbivorous amphipod Ampithoe valida on three dominant macroalgal species of Yundang Lagoon. Acta Ecologica Sinica, 2013, 33(22): 7166-7172 [郑新庆, 黄凌风, 李元超, 等. 啃食性端足类强壮藻钩虾对筼筜湖三种大型海藻的摄食选择性. 生态学报, 2013, 33(22): 7166-7172] |