



2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 山东 青岛 266071;
3. 上海海洋大学海洋科学学院 上海 201306
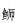 (Seriola lalandi)、绯
(Seriola lalandi)、绯 (Callionymus beniteguri)、少鳞
(Callionymus beniteguri)、少鳞 (Sillago japonica)、黑鲷(Acanthopagrus schlegelii)、短吻红舌鳎(Cynoglossus joyneri)、长蛇鲻(Saurida elongata)等;烟威近岸海域鱼卵与仔稚鱼群落结构年内变化明显,除产卵末期(8—9月)外,各月间种类更替率不低于50%,鱼卵与仔稚鱼群落月间平均相异性不低于73%;海表温度(SST)、海底温度(SBT)、海表盐度(SSS)和深度(DEP)是显著影响4—9月主要鱼种产卵选择的环境因子。研究表明,烟威近岸海域为黄渤海规模较大的鱼类产卵场之一,需在鱼类早期生活史研究与产卵场养护策略制定时得到足够重视。
(Sillago japonica)、黑鲷(Acanthopagrus schlegelii)、短吻红舌鳎(Cynoglossus joyneri)、长蛇鲻(Saurida elongata)等;烟威近岸海域鱼卵与仔稚鱼群落结构年内变化明显,除产卵末期(8—9月)外,各月间种类更替率不低于50%,鱼卵与仔稚鱼群落月间平均相异性不低于73%;海表温度(SST)、海底温度(SBT)、海表盐度(SSS)和深度(DEP)是显著影响4—9月主要鱼种产卵选择的环境因子。研究表明,烟威近岸海域为黄渤海规模较大的鱼类产卵场之一,需在鱼类早期生活史研究与产卵场养护策略制定时得到足够重视。2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot Qingdao National Laboratory for Marine Science and Technology (Qingdao), Qingdao, Shandong 266071, China;
3. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China
渔业种群资源量的变动主要是由其补充量来决定,海洋鱼类通过繁殖、发育和生长使种群因自然死亡和捕捞死亡而导致的减损得到补偿(Hutchings et al, 2004)。产卵场是海洋水产动物早期生活史阶段的重要栖息地,鱼类产卵场的生境条件影响了鱼卵及仔稚鱼的种类、数量和生长发育状况,进而对海洋鱼类补充量产生重要影响(卞晓东等, 2018; Beaugrand et al, 2003)。因此,对产卵场开展鱼卵、仔稚鱼及环境调查,可以从鱼类早期生活史视角探寻渔业资源种类组成及补充状况。
烟威近岸海域鱼类产卵场研究始于1953年新中国首次海洋渔场综合调查“烟、威海区鲐鱼渔场调查”(朱树屏, 1959),该调查探明了烟威渔场地理、水文和浮游生物等环境状况,证实了烟威近岸海域存在重要的鲐(Scomber japonicus)产卵场。1985—1988年,中国水产科学研究院黄海水产研究所于黄海开展了大范围、多航次鱼卵和仔稚鱼调查采集工作,其中于121°~124°30′E、37°~38°N范围内设置39个站位,覆盖整个烟威近岸海域。调查发现,烟威渔场西北部为鱼卵密集分布区,5月盛期主要产卵种类为鳀(Engraulis japonicus)、多鳞
烟威近岸海域为多种经济鱼类亲体生殖洄游进入渤海和其幼体索饵洄游出渤海的重要通道,渔业资源种类多达67种(张玉钦等, 2019)。春季鱼类产卵群体在烟威渔场温盐锋面两侧集群,待机洄游进入渤海或沿岸各海湾产卵;夏季该海域初级生产力亦为黄海较高水平,为各种仔稚鱼的生长发育提供了良好的生境条件(金显仕等, 2005; 张孝威等, 1959)。作为黄渤海鱼类生殖洄游的重要通道和毗邻海域渔业资源补充的重要海域,开展烟威近岸海域产卵场现状调查对摸清水域内主要种类的产卵期、产卵场范围和其补充能力以及洄游性鱼类迁徙路线的研究均具有重要意义,而站位广覆盖的逐月产卵场调查则是剖析烟威近岸海域产卵场现状的首要条件。因此,本研究基于2020年4—9月烟威近岸海域36站位逐月调查采集的鱼卵、仔稚鱼及环境数据,分析该海域鱼卵、仔稚鱼群落结构和数量分布特征及月际变动,获取烟威近岸海域鱼类产卵场现状新认知,为黄渤海鱼类早期资源养护和产卵场保护策略制定提供生态学基础。
1 材料与方法 1.1 数据采集和样品分析2020年4—9月,中国水产科学研究院黄海水产研究所于烟威近岸海域开展了共6航次、每航次36站位的逐月产卵场调查(图 1),调查船为中渔科101/102号。调查时间分别为4月底(4月22日—5月1日)、5月底—6月初(5月30日—6月9日)、6月底(6月25—30日)、7月中旬(7月12—20日)、8月下旬(8月16—25日)和9月中下旬(9月16—25日),调查采集到4—9月期间各月的鱼卵、仔稚幼鱼、海水表底温度、海水表底盐度、水深、叶绿素、浮游动植物及5项营养盐数据。海上调查流程严格遵守《海洋调查规范—海洋生物调查》(GB/T 12763.6-2007)。各航次鱼卵和仔稚鱼采样操作均保持一致,即每个调查站位通过大型浮游生物网(网口内径0.8 m,网长2.7 m,网目孔径0.505 mm)于海水表层水平拖网采集1次,每次以2 n mile/h拖速拖行10 min,并系网口流量计以计算滤水量。浮游动植物则分别采用浅水Ⅱ型网(网口内径0.316 m,网目孔径0.160 mm)和浅水Ⅲ型网(网口内径0.370 m,网目孔径0.070 mm)由海水底层垂直向上采集。表层叶绿素则通过采集500 mL表层海水后,抽滤至玻璃纤维滤膜(孔径0.7 μm),于–20℃冷冻保存。表、底层海水温度和盐度通过美国Seabird 19 plus或德国Sea & Sun Technology CTD 60M温盐深仪采集,将海面向下0.5 m判定为表层海水,海底向上0.5 m判定为底层海水。各采样点的海水深度通过船载声呐获取。

|
图 1 2020年烟威近岸海域产卵场调查站位 Fig.1 Sampling stations of the spawning ground survey in the Yantai-Weihai offshore waters in 2020 |
鱼卵、仔稚鱼和浮游动植物样品采集后,5%海水甲醛溶液固定。鱼卵和仔稚鱼样品全样本分析,体视显微镜下经形态学鉴定并反复核实,参照张仁斋等(1985)、冲山宗雄(2014)和万瑞景等(2016),确认至每一物种单元,并按种类及其发育阶段分别记录个体数,分析过程严格遵守《海洋调查规范—海洋生物调查》(GB/T 12763.6-2007),物种学名引自World Register of Marine Species (WoRMS, https://www.marinespecies.org/)。浮游动植物样品形态学鉴定,样品处理及分析方法严格遵守《海洋调查规范—海洋生物调查》(GB/T 12763.6-2007)。叶绿素滤膜则使用90%丙酮溶液于–20℃冷冻萃取12 h后,利用Turner Designs Trilogy荧光仪测定叶绿素a浓度。
1.2 物种优势度通过Pinkas相对重要性指数(index of relative importance, IRI) (Pinkas, 1971)判断各月份鱼卵、仔稚鱼的优势种(IRI≥1000)、重要种(1000 > IRI≥100)、常见种(100 > IRI > 10)、一般种(10 > IRI > 1)和少见种(IRI < 1) (程济生, 2004),计算公式如下:
| $ \mathrm{IRI}=N \% \times F \% \times 10 $ | (1) |
式中,N%为某种鱼卵或仔稚鱼的数量占当月鱼卵或仔稚鱼总数量的百分比,F%为某种鱼卵或仔稚鱼的站位出现频率。
1.3 物种多样性通过Margalef丰富度指数D (Margalef, 1957)、Shannon-Wiener多样性指数H′ (Shannon, 1948)和Pielou均匀度指数J′ (Pielou, 1969)判断各月份的生物多样性状况,计算公式如下:
| $ D=\frac{S-1}{\ln N} $ | (2) |
| $ \begin{gathered} H^{\prime}=-\sum\limits_{i=1}^S \frac{N_i}{N} \ln \frac{N_i}{N} \end{gathered} $ | (3) |
| $ J^{\prime}=\frac{H^{\prime}}{\ln S} $ | (4) |
式中,S为当月鱼卵或仔稚鱼物种数量;Ni为当月物种i的鱼卵或仔稚鱼数量;N为当月所有物种的鱼卵或仔稚鱼总数量。
1.4 群落结构月度变动分析种类更替率(E)采用各月鱼卵与仔稚鱼物种的并集进行计算,公式为(卞晓东等, 2010):
| $E=\frac{A}{A+B} $ | (5) |
式中,A为2个月间种类增加数与减少数之和;B为2个月间相同种类数。
构建Bray-Curtis相似度矩阵,对鱼卵、仔稚鱼丰度数据按照月份尺度进行聚类分析(cluster analysis)及非度量多维标度分析(non-metric multi-dimensional scaling analysis, NMDS)以对群落结构进行研究。通过相似性分析(analysis of similarities, ANOSIM)研究组间差异显著性,通过胁强系数(stress)检验NMDS结果可靠性,若Stress < 0.05,说明结果具有很好的代表性(Khalaf et al, 2002; Crona et al, 2006)。将鱼卵及仔稚鱼丰度(即单位滤水量采集到的鱼卵数或仔稚鱼数, Abu)作为Cluster及NMDS月份–物种二维矩阵元素值,计算公式为:
| $\mathrm{Abu}=\frac{N}{R \times q \times \pi \times r} $ | (6) |
式中,N为鱼卵数量(ind.);R为网口流量计转数差值(round);q为网口流量计的距离转数比(m/round);r为大型浮游生物水平拖网的网口半径(m)。
对2020年4—9月鱼卵与仔稚鱼种类进行相邻月间相似性百分比分析(similarity percentage analysis, SIMPER),结合月间种类更替率与各物种的平均相异性贡献率,反映相邻月间群落结构更替情况及主要分歧种(平均相异性贡献率之和达90%的物种);对Cluster和NMDS分出的鱼卵与仔稚鱼种类各月份组进行相似性分析,通过各物种的平均相似性贡献率反映不同月份组的主要特征种(平均相似性贡献率之和达90%的物种)。
1.5 环境因子分析冗余分析(redundancy analysis, RDA)和典范对应分析(canonical correspondence analysis, CCA)是识别多物种变量与多环境因子变量间相关性的有效方法(潘玉龙等, 2019; 梁姗姗等, 2021)。由于2020年烟威近岸海域调查中,4月和9月鱼卵和仔稚鱼种类、数量、丰度及出现频率均较低,因此,选择主要产卵季节5—8月出现频率≥5%的鱼卵、仔稚鱼种类构建物种变量矩阵(温超男等, 2020),并对其丰度(Abu)进行[log(x+1)]对数转换以降低极端值影响(樊紫薇等, 2020)。采用物种变量矩阵中各站位对应的环境数据构建环境变量矩阵,包括海表温度(SST)、海底温度(SBT)、海表盐度(SSS)、海底盐度(SBT)、深度(DEP)、海表叶绿素a浓度(SCHLA)、浮游动物总数(ZP)和浮游植物总数(PP)共8个环境变量。
对物种变量矩阵通过除趋势对应分析(detrended correspondence analysis, DCA)结果中第1排序轴的梯度长度判断采用RDA或CCA,若第1排序轴的梯度长度 > 4.0,应采用CCA分析;若梯度长度处于[3.0, 4.0]区间内,CCA和RDA均可使用;若梯度长度 < 3.0,RDA分析是更好的选择(Ter Braak, 1986)。通过蒙特卡洛置换检验(Monte Carlo permutation tests)判断环境变量显著性,保留P < 0.05的环境变量构建最终模型,判断环境因子对鱼卵及仔稚鱼优势种和重要种的影响。
1.6 数据分析及可视化工具使用ArcGIS 10.2对2020年4—9月烟威近岸海表温度进行普通克里金空间插值和绘图,使用Excel 2016计算物种优势度、多样性指数和更替率,通过Primer 6进行群落结构分析,通过R 4.0.2和Canoco 5分析物种与环境因子的相关性。
2 结果 2.1 鱼卵及仔稚鱼时空分布2020年4—9月鱼卵的站位分布情况如图 2所示。4—9月共采集鱼卵337 038粒,其中,4月底采集46粒,出现频率为33.33%,站均1.28粒,平均丰度为3.09×10–3 ind./m3,鱼卵分布于烟台蓬莱–牟平北部海域、威海鸡鸣岛北部近岸海域以及桑沟湾东部外海,最大丰度为0.04 ind./m3;5月底—6月初采集鱼卵287 844粒,出现频率为94.44%,站均7 995.67粒,平均丰度为35.40 ind./m3,烟台蓬莱市–威海鸡鸣岛北部大面海域分布着大量鱼卵,以烟台牟平–威海刘公岛北部海域的站卵量水平最高,最大丰度为498.15 ind./m3;6月底采集鱼卵43 542粒,出现频率为100%,站均1 209.5粒,平均丰度为3.99 ind./m3,鱼卵集中分布在烟台牟平–威海鸡鸣岛北部离岸海域,最大丰度为29.75 ind./m3;7月中旬采集4751粒鱼卵,出现频率为94.44%,站均131.97粒,平均丰度为0.69 ind./m3,烟台牟平–威海鸡鸣岛北部离岸海域和荣成湾–桑沟湾东部近岸海域的站卵量水平较高,最大丰度为11.54 ind./m3;8月下旬采集641粒鱼卵,出现频率为77.78%,站均17.81粒,平均丰度为0.05 ind./m3,烟威交界–桑沟湾外海具有较高的鱼卵出现频率和站卵量水平,最大丰度为0.71 ind./m3;9月中下旬采集鱼卵214粒,出现频率为33.33%,站均5.94粒,平均丰度为0.01 ind./m3,鱼卵主要分布在烟台牟平–威海鸡鸣岛北部外海和成山角–荣成湾近岸海域,最大丰度为0.22 ind./m3。

|
图 2 2020年4—9月烟威近岸海域鱼卵数量分布及海表温度月度变动 Fig.2 Monthly distribution patterns of fish eggs and sea surface temperature in the Yantai-Weihai offshore waters from April to September, in 2020 |
2020年4—9月仔稚鱼的站位分布情况见图 3。4—9月共采集仔稚鱼2122尾,其中,4月底采集到181尾,出现频率为47.22%,站均5.03尾,平均丰度为6.12×10–3 ind./m3,仔稚鱼主要分布在威海刘公岛–荣成湾近岸海域,最大丰度为0.18 ind./m3;5月底—6月初采集到271尾,出现频率为47.22%,站均7.53尾,平均丰度为0.04 ind./m3,仔稚鱼主要分布在烟台莱山区北部近岸海域和威海荣成湾东部近岸海域,最大尾数为163尾,最大丰度为0.96 ind./m3;6月底采集到186尾,出现频率为77.78%,站均5.17尾,平均丰度为0.02 ind./m3,仔稚鱼分布较均匀,其中,烟台牟平–威海刘公岛北部海域仔稚鱼数量较多,最大丰度为0.12 ind./m3;7月中旬采集1323尾,出现频率为72.22%,站均36.75尾,平均丰度为0.11 ind./m3,烟台牟平北部外海和近岸海域、威海刘公岛近岸海域以及鸡鸣岛北部外海分布着大量仔稚鱼,最大丰度为1.50 ind./m3;8月下旬采集92尾,出现频率为61.11%,站均2.56尾,平均丰度为7.63× 10–3 ind./m3,仔稚鱼分布较均匀,最大丰度为0.05 ind./m3;9月中下旬采集69尾,出现频率为58.33%,站均1.92尾,平均丰度为5.66×10–3 ind./m3,烟台套子湾北部近岸海域、威海刘公岛北部海域和荣成湾东部海域仔稚鱼数量较多,最大丰度为0.02 ind./m3。

|
图 3 2020年4—9月烟威近岸海域仔稚鱼数量分布及海表温度月度变动 Fig.3 Monthly distribution patterns of fish larvae and sea surface temperature in the Yantai-Weihai offshore waters from April to September, in 2020 |
2020年4—9月共采集33种鱼卵,1种未定种,32种鉴定到种,隶属6目21科。已鉴定种类中,鲈形目(Perciformes) 21种,鲽形目(Pleuronectiformes) 5种,鲱形目(Clupeiformes) 4种,灯笼鱼目(Myctophiformes)和鲻形目(Mugiliformes)各1种;共采集仔稚鱼28种,隶属8目22科,其中,鲈形目16种,颌针鱼目(Beloniformes)和鲻形目各3种,海龙目(Syngnathiformes) 2种,鲱形目、银汉鱼目(Atheriniformes)、灯笼鱼目和鲽形目各1种。从时间上看,4月底采集到4种鱼卵和4种仔稚鱼,5月底—6月初采集到21种鱼卵和6种仔稚鱼,6月底采集到19种鱼卵和13种仔稚鱼,7月中旬采集到13种鱼卵和12种仔稚鱼,8月下旬采集到11种鱼卵和10种仔稚鱼,9月中下旬采集到9种鱼卵和8种仔稚鱼,其中,5月底—6月初采集到的鱼卵种类数最多,6月底采集到的仔稚鱼种类最多,鳀的鱼卵和仔稚鱼均具有最高的月份出现频率(表 1)。
|
|
表 1 2020年4—9月烟威近岸海域鱼卵及仔稚鱼种类组成 Tab.1 Species composition of fish eggs and larvae in the Yantai-Weihai offshore waters from April to September, in 2020 |
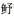 科Sphyraenidae
科Sphyraenidae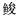 Planiliza haematocheilus
Planiliza haematocheilus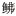 科Triglidae
科Triglidae2020年4月底,烟威近岸海域鱼卵优势种为尖吻黄盖鲽(Pseudopleuronectes herzensteini),重要种为鲐(Scomber japonicus);仔稚鱼优势种为玉筋鱼(Ammodytes personatus),重要种为方氏云鳚(Pholis fangi)。5月底—6月初,鱼卵优势种为鳀和鲐,无重要种;仔稚鱼优势种为许氏平鲉(Sebastes schlegelii),重要种为鳀。6月底,鱼卵优势种为鳀,重要种为鲐;仔稚鱼优势种为鳀,重要种为沙氏下
2020年4—9月,烟威近岸海域鱼卵及仔稚鱼的群落多样性有明显的月度变动(图 4)。鱼卵丰富度指数(D)在4月最低,5—9月均较高,其中6月最高;鱼卵多样性指数(H′)在6月最低,7—9月均较高,其中8月最高;鱼卵均匀度指数(J′)在6月最低,4月最高。仔稚鱼丰富度指数(D)在4月和5月水平较低,6—9月较高,其中6月最高;仔稚鱼多样性指数(H′)在4月、5月和7月较低,在6月、8月和9月较高,其中8月最高;仔稚鱼均匀度指数(J′)在8月最高,7月则明显低于其他月份。

|
图 4 2020年4—9月烟威近岸海域鱼卵(a)及仔稚鱼(b)多样性指数
Fig.4 Diversity index of fish eggs (a) and larvae (b) in the Yantai-Weihai offshore waters from April to September, in 2020
 :Margalef丰富度指数D; :Margalef丰富度指数D; :Shannon-Wiener多样性指数H′; :Shannon-Wiener多样性指数H′; 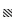 : Pielou均匀度指数J′
: Margalef richness index D; : Shannon-Wiener diversity index H′; : Pielou evenness index J′ : Pielou均匀度指数J′
: Margalef richness index D; : Shannon-Wiener diversity index H′; : Pielou evenness index J′
|
2020年4—9月共采集到52种鱼卵和仔稚鱼,4—5月鱼卵及仔稚鱼种类更替率为90.00%,鱼卵群落平均相异性为99.98%,仔稚鱼群落平均相异性为99.68%;5—6月种类更替率为51.35%,鱼卵群落平均相异性为80.65%,仔稚鱼群落平均相异性为81.37%;6—7月种类更替率为50.00%,鱼卵群落平均相异性为73.34%,仔稚鱼群落平均相异性为78.79%;7—8月种类更替率为53.57%,鱼卵群落平均相异性为87.83%,仔稚鱼群落平均相异性为93.47%;8—9月种类更替率为47.82%,鱼卵群落平均相异性为81.77%,仔稚鱼群落平均相异性为59.03% (图 5)。从图 5可以看出,4—5月具有最高的鱼卵及仔稚鱼种类更替率、鱼卵群落平均相异性和仔稚鱼群落平均相异性,7—8月次之。8—9月种类更替率最低,6—7月鱼卵群落平均相异性最低,8—9月仔稚鱼群落平均相异性最低。

|
图 5 月间鱼卵及仔稚鱼种类更替率及平均相异性
Fig.5 Monthly species turnover rates and average dissimilarities of fish eggs and larvae
 :月间种类更替率; :月间种类更替率;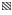 :鱼卵群落平均相异性; :鱼卵群落平均相异性; :仔稚鱼群落平均相异性
: Monthly species turnover rates; : Average dissimilarities of fish egg communities; : Average dissimilarities of fish larva communities :仔稚鱼群落平均相异性
: Monthly species turnover rates; : Average dissimilarities of fish egg communities; : Average dissimilarities of fish larva communities
|
4—5月鱼卵群落主要分歧种为鳀和鲐,仔稚鱼群落主要分歧种为许氏平鲉、褐菖鲉(Sebastiscus marmoratus)、玉筋鱼和鳀;5—6月鱼卵群落主要分歧种为鳀和鲐,仔稚鱼群落主要分歧种为许氏平鲉、褐菖鲉、鳀、沙氏下
|
|
表 2 2020年4—9月烟威近岸海域鱼卵和仔稚鱼优势种及重要种 Tab.2 Dominant and important species of eggs and larvae in the Yantai-Weihai offshore waters from April to September, in 2020 |
|
|
表 3 月间鱼卵及仔稚鱼平均相异性贡献率之和占90%以上的分歧种 Tab.3 The discriminating species that contribute more than 90% to the monthly average dissimilarity of fish eggs and larvae |
选取2020年5—8月鱼卵和仔稚鱼优势种、重要种及其他出现频率较高且具较大经济价值或食物网研究价值的物种构建物种变量矩阵,该矩阵由鳀、鲐、蓝点马鲛(Scomberomorus niphonius)、黄条
|
|
表 4 聚类组内(图 6中A~D组)鱼卵及仔稚鱼平均相似性贡献率之和占90%以上的特征种 Tab.4 The tipical species that contribute more than 90% of the average similarity in cluster groups (from A to D in Fig. 6) of fish eggs and larvae |
|
|
表 5 RDA分析中显著环境变量的统计学参数 Tab.5 Statistical parameters of significant environmental variables in RDA |

|
图 6 2020年4—9月鱼卵(a)和仔稚鱼(b)种类聚类图及NMDS排序图 Fig.6 Monthly scale clustering diagram and NMDS ordination diagram of fish eggs (a) and larvae (b) from April to September, in 2020 |

|
图 7 鱼卵及仔稚鱼种类与环境因子间RDA排序图
Fig.7 RDA ordination of fish egg and larva taxas with environmental factors
各编号代表的鱼类物种如表 1所示; :环境因子; :环境因子; :鱼卵物种; :鱼卵物种; :仔稚鱼物种
The fish species names corresponding to the numbers are shown in Tab. 1; : Environmental factors;
: Fish egg taxas; : Fish larva taxas :仔稚鱼物种
The fish species names corresponding to the numbers are shown in Tab. 1; : Environmental factors;
: Fish egg taxas; : Fish larva taxas
|
与1985—1988年调查(万瑞景等, 1998)结果相似,2020年4—9月烟威近岸海域鱼卵和仔稚鱼的数量有明显时空变化,产卵盛期均在5—6月,仔稚鱼盛期在7月。1985—1988年调查结果显示,鱼卵密集中心位于烟威渔场偏西部海域,9月后由烟威近岸和海洋岛产卵场游出并进行越冬洄游的鱼类早期资源在烟威渔场东部海域聚集(万瑞景等, 1998)。2020年4—9月调查结果显示,鱼卵密集中心位于烟台套子湾至威海鸡鸣岛北部海域,仔稚鱼则主要分布在烟台牟平–威海鸡鸣岛北部海域;由于2020年调查站位未覆盖123°E以东海域,因此,本研究未能针对该海域仔稚鱼在9月的分布情况进行相关讨论。4月底,烟威近岸海域盐度较高但水温较低,尚未达到绝大多数暖温性鱼类的产卵水温要求,仅在烟台牟平以西、威海鸡鸣岛近岸和桑沟湾近岸海域采集少量鱼卵,在威海刘公岛–荣成湾近岸海域采集部分仔稚鱼。5月底—6月初,海域盐度稍有下降但整体水平较高,且随着水温持续上升,多数鱼类进入产卵期,产卵场范围覆盖整个山东半岛北部黄海近岸,平均水温最高的烟台套子湾–牟平北部海域也是卵量最高值区域;同期威海东部近岸海域仔稚鱼尾数高值区扩大,且烟台北部海域仔稚鱼数量水平有所提升。6月底,水温进一步升高,产卵盛期尚未结束,主要产卵场范围缩小并移至烟台牟平至威海鸡鸣岛北部离岸海域;仔稚鱼密度仅与4月底相当,但出现频率提升至全年最高水平,高值区由威海东部近岸移至烟台牟平–威海刘公岛北部海域。7月,烟台套子湾以东海域的水温已高于20.4℃,但海表盐度明显低于4—6月,鱼类产卵盛期结束,鱼卵出现频率虽仍保持在90%以上,但鱼卵数量超过1000粒的站位仅剩1站;仔稚鱼数量达到全调查月份最高水平,鳀仔稚鱼开始大量出现,分布高值区由威海东部近岸移至烟台牟平–威海刘公岛北部海域。8月,烟威北部海域的水温升至24.1℃以上,鱼类亲体的产卵种类和规模骤减,除鳀和短吻红舌鳎外,仅有少量暖水性鱼类产卵繁殖,平均站卵量和鱼卵出现频率均下降,卵量高值区东移至威海北部、东部近岸海域;仔稚鱼出现频率仍明显高于4月底和5月底—6月初,但仔稚鱼密度明显降低。9月起海域水温开始下降,烟威近岸海域鱼卵数量和出现频率骤降,卵量高值区移至威海刘公岛北部离岸海域;仔稚鱼数量降至全调查月份次最低水平。
3.2 烟威近岸海域鱼卵及仔稚鱼群落结构及月度更替中的特征种和分歧种2020年4—9月,烟威近岸海域共采集到鱼类早期资源种类52种,包括33种鱼类的337 038粒鱼卵和28种鱼类的2122尾仔稚鱼。鱼卵与仔稚鱼群落结构年内变化明显,除产卵末期8—9月外,各月间种类更替率不低于50%,鱼卵与仔稚鱼群落月间平均相异性不低于73%。烟威近岸海域温盐环境的显著变化是鱼类早期资源群落结构更替的主要原因。4月底,烟威近岸海域海水温度较低,早期资源种类以玉筋鱼、尖吻黄盖鲽、方氏云鳚、许氏平鲉等冷温性鱼类为主;5月底—6月初,该海域平均表温已高于16℃,海水表底层盐度的平均值高于32.4,且在南北方向上梯度变化明显(北高南低),为鳀、鲐、蓝点马鲛、黄条
2020年4—9月,烟威近岸海域共出现33种鱼卵,鳀在5—9月的3个群落中均为鱼卵特征种,鲐是4月群落和5月群落鱼卵特征种,尖吻黄盖鲽是4月群落鱼卵特征种,绯
2020年4—9月,烟威近岸海域共出现28种仔稚鱼,鳀在5—9月的3个群落中均为仔稚鱼特征种,方氏云鳚和玉筋鱼是4月群落仔稚鱼特征种,许氏平鲉和褐菖鲉是5月群落仔稚鱼特征种,纹缟
烟威近岸海域环境因子在4—9月变动明显,调查海域海表温度(SST)范围为8.27~26.05℃,海底温度(SBT)范围为8.01~24.52℃,海表盐度(SSS)范围为30.87~33.27,海水深度范围为4.4~82.0 m,海表温度、海底温度、海表盐度与深度的时空变动显著影响鱼类产卵选择。各鱼种产卵月份、主要产卵区域及适宜产卵生境如表 6所示。
|
|
表 6 主要鱼种产卵月份、主要产卵区域及适宜产卵生境 Tab.6 Spawning month, major spawning areas and suitable spawning habitats of main species |
鳀为暖温性中上层洄游鱼类,是海洋鱼类重要的饵料生物(唐启升等, 2005),其鱼卵在黄渤海(万瑞景等, 2008; 阮洪超, 1984)各产卵场中占较大比重,水温上升是驱动鳀产卵洄游并在4月下旬抵达黄海北部和渤海的重要条件(卞晓东等, 2022b; 金显仕等, 2005)。鳀在烟威近岸海域产卵期为4—9月,其鱼卵在5—9月鱼卵群落中均表现最高优势度,主要产卵期为5—6月,鳀适宜产卵生境为低温高盐海域,鳀卵主要分布于海表盐度32.0~33.0、海表温度16.4~21.7℃、海底温度8.5~15.7℃、深度11.0~43.2 m的烟威北部海域,水温和盐度对鳀卵分布影响较大。
鲐为暖水性中上层洄游鱼类,在我国黄海、东海有集中分布,具有重要的经济价值(严利平等, 2012)。20世纪50年代,烟威外海是重要的鲐产卵场(朱树屏, 1959),但近年来鲐资源量波动较大(陈大刚, 1991),其补充量分布规律及资源评估研究受到广泛关注(李曰嵩等, 2018; Takasuka et al, 2008)。鲐于4—7月在烟威近岸海域产卵,主要产卵期为5—6月,其鱼卵为5月优势种和4月、6月重要种,鲐亲鱼适宜在低温高盐海洋环境产卵,鲐卵主要分布于海表盐度32.0~33.0、海表温度16.4~21.7℃、海底温度8.5~16.2℃、深度26.5~43.2 m的烟台牟平–威海刘公岛北部海域中。
蓝点马鲛为洄游性中上层鱼类,具有重要的经济价值(邱盛尧等, 1993; 祖凯伟等, 2019),适宜产卵生境为低温高盐海洋环境(卞晓东等, 2022b)。5月底—6月初于烟威近岸海域采集到大量蓝点马鲛卵,不过产卵期较短,其鱼卵在6月底之后基本消失。水温及盐度对蓝点马鲛鱼卵丰度影响明显,低温高盐海洋环境适宜蓝点马鲛产卵,其鱼卵主要分布于海表盐度32.0~32.6、海表温度16.9~19.8℃、海底温度8.6~16.2℃、深度10.6~43.2 m的烟台莱山区–威海刘公岛北部海域。值得一提的是,同为大尺度洄游性中上层鱼类的鳀、蓝点马鲛和鲐的产卵盛期与密集中心高度重合,而鳀在蓝点马鲛和鲐的食物组成中均占重要地位(张波, 2018; 张孝威等, 1959),因此,在饵料层面上,鳀群分布对鲐和蓝点马鲛产卵选择影响深刻,这一点也在烟威近岸海域鲐产卵区域影响因子研究(张雨轩等, 2022)及黄、渤海蓝点马鲛产卵场分布规律研究(韦晟, 1980)中得到印证。
高眼鲽为冷温性底层鱼类(万瑞景等, 1998),是黄海渔业生态系统中的主要种类(滕广亮等, 2018),其越冬场位于石岛东部水域,主要产卵场位于山东南部、山东北部、海洋岛等海域。烟威近岸海域高眼鲽产卵期由5月持续至9月,5月底—6月初为其产卵盛期,之后仅零星分布,低温高盐环境适宜高眼鲽产卵,其鱼卵主要分布于海表盐度32.2~33.0、海表温度16.9~19.1℃、海底温度8.6~13.5℃、深度19.8~43.2 m的烟台牟平–威海刘公岛北部离岸海域。
黄条
少鳞
黑鲷为暖温性底层鱼类,喜栖息于近岸岩礁海区、内湾及咸淡水水域(陈大刚等, 2015),具极高经济价值,是我国重要的增殖放流对象(李冬辰等, 2021)。烟威近岸海域黑鲷产卵期为5月底—6月初,其鱼卵分布于海表盐度32.0~32.6、海表温度16.4~19.6℃、海底温度13.5~16.9℃、深度10.4~23.7 m的烟台蓬莱–莱山区北部近岸海域,尤以烟台莱山区北部近岸1站鱼卵数量最高,占该航次黑鲷鱼卵总数的92.60%。温度和水深是影响黑鲷产卵选择的显著因子,其适宜产卵生境为低温浅水近岸海域。
长蛇鲻为暖温性底层鱼类,是重要的经济鱼种(陈大刚, 1991)。烟威近岸海域长蛇鲻产卵期较长,为5—8月,6—7月为主要产卵期,适宜在底温较高、盐度较低的较浅海水中产卵,其鱼卵主要分布在海表盐度31.0~32.6、海表温度19.3~21.9℃、海底温度15.3~20.6℃、深度16.8~30.4 m的烟台蓬莱–莱山区北部海域。
尖吻黄盖鲽为近海底层洄游性鱼类,具有一定的经济价值,其亲鱼于每年3月由石岛外海深水区越冬场分2支北上产卵洄游,一支4月游抵渤海海峡,另一支5月游至海洋岛附近产卵(陈大刚, 1991; 毕庶万等, 1993)。4月底已有尖吻黄盖鲽于烟台蓬莱–烟威交界北部海域产卵,之后不再出现,6月下旬发现其仔稚鱼,其鱼卵主要分布于海表盐度32.3~33.0、海表温度9.6~12.2℃、海底温度8.4~11.2℃、深度8.4~24.2 m的烟台蓬莱–牟平以北低温高盐浅水海域。
3.4 烟威近岸海域硬骨鱼类产卵场重要性中国水产科学研究院黄海水产研究所于1985—1988年对黄海硬骨鱼类产卵场进行综合调查(万瑞景等, 1998),该调查与本调查相比,网具规格、取样方法和每站拖网时间基本一致,也基本覆盖了烟威近岸海域硬骨鱼类主要产卵期。1986年5月15日—5月29日黄海大面86站调查共采集14种硬骨鱼类的299 592粒鱼卵,平均资源密度为3 524.6 ind./haul,密度最高的海州湾渔场和连青石渔场西南部海域有4站鱼卵数量高达2.8~16.6万ind./haul,为黄海主要中心产卵场之一,而烟威渔场西北部密集区的鱼卵密度仅为500~4000 ind./haul;1985年6月25日—7月12日黄海北段海域63站调查共采集15种鱼类的138 942粒鱼卵,作为最大密集分布区的石岛渔场东南部至海洋岛渔场东南部海域资源密度均在1000 ind./haul以上,最大2站达18 738和21 112 ind./haul,烟威渔场西部和中部形成2个1000 ind./haul以上的密集分布区,最高密度分别达30 000和7231 ind./haul (万瑞景等, 1998)。与该调查结果不同,2020年5月底—6月初,本调查于烟威近岸海域36站共采集到21种鱼类的287 844粒鱼卵,平均资源密度为7995.67 ind./haul,卵量最高的烟台牟平近岸站位采集到12万余粒鱼卵,另有4站鱼卵数量为10 000~40 000粒,远高于1985—1986年调查中烟威渔场的资源密度水平。当前调查,烟威近岸海域除鳀和鲐等主要种类外,仍出现20余种产卵期短暂或出现频率与数量占比较低的鱼类早期资源,其中不乏带鱼、蓝点马鲛和小黄鱼(Larimichthys polyactis)等黄渤海重要经济鱼种,符合其“过路渔场”属性。不过,本研究在1986—1988年调查(万瑞景等, 1998)未覆盖到的37°30′N以南的烟威近岸海域发现大量鳀卵和鲐卵;此外,黄条
渤海莱州湾为黄渤海众多渔业生物关键栖息地,是黄渤海渔业生物主要产卵场、育幼场和渤海多种渔业的传统渔场(Jin et al, 2013),该海域鱼卵平均资源密度季节变化规律与烟威近岸海域相同,均为升温期(春季、春夏季)最高,夏季急剧下降,秋季仅有零星鱼卵分布(卞晓东等, 2022a);不过,近年莱州湾春季至春夏季(5—6月)产卵盛期鱼卵平均资源密度(春季1 473.61 ind./haul,春夏季146~983.14 ind./haul)及种类数量(春季16种,春夏季18种)均明显低于烟威近岸海域同期水平,作为烟威近岸海域5—6月重要产卵鱼种—鲐、高眼鲽、黄条
本研究基于2020年4—9月逐月调查数据,对山东半岛北部烟威近岸海域鱼类产卵场现状进行了系统阐述。结果显示,2020年4—9月,烟威近岸海域鱼类产卵场分布着大量鱼类早期资源群体,有52种鱼类的鱼卵或仔稚鱼在该产卵场内出现,产卵盛期为5—6月,仔稚鱼盛期为7月,主要产卵场分布在烟台套子湾至威海鸡鸣岛北部海域,仔稚鱼主要生长发育场所位于烟台牟平–威海鸡鸣岛北部海域。鱼类早期资源群落结构的月际变化明显,4月多为冷温性鱼类,主要产卵期内以海水中上层暖温性、暖水性鱼类为主导,产卵末期则多为近岸浅水或岩礁性鱼类,海水温盐环境的时空差异是鱼类早期资源数量变动及群落结构更替的主要原因,7月中旬之后的低盐度水平是产卵盛期持续性的重要限制因子。综合来看,烟威近岸海域不仅是带鱼、小黄鱼和蓝点马鲛等多种重要经济鱼类的“过路渔场”,也是鳀、鲐、高眼鲽和黄条
BEAUGRAND G, BRANDER K M, LINDLEY J A, et al. Plankton effect on cod recruitment in the North Sea. Nature, 2003, 426(6967): 661-664 DOI:10.1038/nature02164 |
BI S W, ZHENG Z H, FENG D G, et al. Preliminary study on the hatching of fertilized eggs and the rearing oflarvae and juveniles of Pseuodopleuronectes herzensteini (Jordan & Snyder). Transactions of Oceanology and Limnology, 1993(4): 75-80 [毕庶万, 郑振虎, 冯敦国, 等. 赫氏黄盖鲽受精卵孵化与仔幼鱼培育初步研究. 海洋湖沼通报, 1993(4): 75-80] |
BIAN X D. Investigation report on the spawning grounds in offshore waters of the Bohai Sea and Yellow Sea from 2014 to 2018 (unpublished). Yellow Sea Fishery Research Institute Scientific Report, 2018 [卞晓东. 2014–2018年黄渤海近岸产卵场调查报告(未公开发表). 黄海水产研究所调查报告, 2018]
|
BIAN X D, WAN R J, JIN X S, et al. Early life resources assemblage structure and succession to the marine Osteichthyes in the Laizhou Bay of Bohai Sea. Journal of Fisheries of China, 2022a, 46(1): 51-72 [卞晓东, 万瑞景, 金显仕, 等. 渤海莱州湾硬骨鱼类早期资源群落结构及演变. 水产学报, 2022a, 46(1): 51-72] |
BIAN X D, WAN R J, SHAN X J, et al. Preliminary analysis on recruitment variation and the exogenous driving factors to early life stages of small pelagic fishes in the Laizhou Bay. Journal of Fishery Sciences of China, 2022b, 29(3): 446-468 [卞晓东, 万瑞景, 单秀娟, 等. 莱州湾中上层小型鱼类早期补充量动态及其外在驱动因素. 中国水产科学, 2022b, 29(3): 446-468] |
BIAN X D, WAN R J, JIN X S, et al. Ichthyoplankton succession and assemblage structure in the Bohai Sea during the past 30 years since the 1980s. Progress in Fishery Sciences, 2018, 39(2): 1-15 [卞晓东, 万瑞景, 金显仕, 等. 近30年渤海鱼类种群早期补充群体群聚特性和结构更替. 渔业科学进展, 2018, 39(2): 1-15] |
BIAN X D, ZHANG X M, GAO T X, et al. Category composition and distributional patterns of ichthyoplankton in the Yellow River estuary during spring and summer 2007. Journal of Fishery Sciences of China, 2010, 17(4): 815-827 [卞晓东, 张秀梅, 高天翔, 等. 2007年春、夏季黄河口海域鱼卵、仔稚鱼种类组成与数量分布. 中国水产科学, 2010, 17(4): 815-827] |
CHAI X L, LI X X, LU R M, et al. Karyotype analysis of the yellowtail kingfish Seriola lalandi lalandi (Perciformes: Carangidae) from South Australia. Aquculture Research, 2009, 40(15): 1735-1741 DOI:10.1111/j.1365-2109.2009.02278.x |
CHEN D G. Fishery Ecology in the Yellow Sea and Bohai Sea. Beijing: Ocean Press, 1991: 326-330 [陈大刚. 黄渤海渔业生态学. 北京: 海洋出版社, 1991: 183-442]
|
CHENG J S. Ecological environment and biotic community in the coastal waters of the Yellow Sea and Bohai Sea. Qingdao: China Ocean University Press, 2004: 244-260 [程济生. 黄渤海近岸水域生态环境与生物群落. 青岛: 中国海洋大学出版社, 2004: 244-260]
|
CRONA B I, HOLMGREN S, RÖNNBÄCK P. Re-establishment of epibiotic communities in reforested mangroves of Gazi Bay, Kenya. Wetlands Ecology and Management, 2006, 14(6): 527-538 DOI:10.1007/s11273-006-9005-7 |
FAN Z W, JIANG R J, LI Z, et al. Community structure of ichthyoplankton and its relationship with environmental factors in Zhongjieshan Islands, China. Acta Ecologica Sinica, 2020, 40(13): 4392-4403 [樊紫薇, 蒋日进, 李哲, 等. 中街山列岛海域鱼卵、仔稚鱼群落结构特征及其与环境因子的关系. 生态学报, 2020, 40(13): 4392-4403] |
HUTCHINGS J A, REYNOLDS J D. Marine fish population collapses: Consequences for recovery and extinction risk. BioScience, 2004, 54(4): 297-309 DOI:10.1641/0006-3568(2004)054[0297:MFPCCF]2.0.CO;2 |
JIN X S, SHAN X J, LI X S, et al. Long-term changes in the fishery ecosystem structure of Laizhou Bay, China. Science China Earth Sciences, 2013, 56(3): 366-374 DOI:10.1007/s11430-012-4528-7 |
JIN X S, ZHAO X Y, MENG T X, et al. The Yellow Sea and Bohai Sea biological resources and habitats. Beijing: Science Press, 2005: 263-264 [金显仕, 赵宪勇, 孟田湘, 等. 黄渤海生物资源与栖息环境. 北京: 科学出版社, 2005: 263-264]
|
KHALAF M A, KOCHZIUS M. Changes in trophic community structure of shore fishes at an industrial site in the Gulf of Aqaba, Red Sea. Marine Ecology Progress Series, 2002, 239: 287-299 DOI:10.3354/meps239287 |
LI D C, ZHANG F. Ingestion of large jellyfish Aurelia coerulea by black sea bream Acanthopagrus schlegelii. Oceanologia et Limnologia Sinica, 2021, 52(6): 1430-1437 [李冬辰, 张芳. 黑鲷(Acanthopagrus schlegelii)对海月水母(Aurelia coerulea)的摄食研究. 海洋与湖沼, 2021, 52(6): 1430-1437] |
LI Y S, BAI S L, PAN L Z, et al. An individual model based simulation study on impact of water temperature fluctuation on recruitment of chub makerel (Scomber japonicus) in the East China Sea. Transactions of Oceanology and Limnology, 2018(6): 118-124 [李曰嵩, 白松麟, 潘灵芝, 等. 基于个体模型水温变动对东海鲐鱼补充量影响模拟研究. 海洋湖沼通报, 2018(6): 118-124] |
LIANG S S, WANG J J, HUANG J S, et al. Ecological vulnerability assessment of coral community under the impact of multiple environmental factors. Ecology and Environmental Sciences, 2021, 30(12): 2360-2369 [梁姗姗, 王建佳, 黄锦树, 等. 近岸多源环境因素影响下珊瑚群落的生态脆弱性评价研究. 生态环境学报, 2021, 30(12): 2360-2369] |
LIU X S, WU J N, HAN G Z, et al. Investigation and division of fisheries resource of Yellow and Bohai Sea. Beijing: Ocean Press, 1990: 78-266 [刘效舜, 吴敬南, 韩光祖, 等. 黄渤海区渔业资源调查与区划. 北京: 海洋出版社, 1990: 78-266]
|
MARGALEF D R. Information theory in ecology. Memorias de la Real Academica de ciencias y artes de Barcelona, 1957, 32: 374-559 |
OKIYAMA M. An atlas of the early stage stage fishes in Japan. second edition. Tokyo: Tokai University Press, 2014 [冲山宗雄. 日本产稚鱼图鉴. 第二版. 东京: 东海大学出版会, 2014]
|
PAN Y L, LIU X, SHA J J, et al. Influence of environmental factors on phytoplanton community structure and its relationship with coastal aquaculture in the waters adjacent to Rongcheng. Progress in Fishery Sciences, 2019, 40(5): 26-33 [潘玉龙, 刘潇, 沙婧婧, 等. 荣成近岸养殖海域浮游植物群落结构及与环境因子的关系. 渔业科学进展, 2019, 40(5): 26-33] |
IELOU E C. An introduction to mathematical ecology. New York: Wiley-Interscience, 1969
|
PINKAS L. Food habits of albacore, bluefin tuna and bonito in California waters. California Department of Fish and Game, Fish Bulletin, 1971, 152: 1-105 |
QIU S Y, YE M Z. The characteristics of growth for Spanish mackerel underyearing in the Yellow Sea and Bohai Sea. Journal of Fisheries of China, 1993, 17(1): 14-23 [邱盛尧, 叶懋中. 黄渤海蓝点马鲛当年幼鱼的生长特性. 水产学报, 1993, 17(1): 14-23] |
RUAN H C. Studies on the eggs and larvae of Engraulis japonicus. Studia Marina Sinica, 1984, 22: 29-56 [阮洪超. 鳀鱼卵子和仔稚鱼的形态发育及其在黄海、渤海的分布. 海洋科学集刊, 1984, 22: 29-56] |
SHANNON C E. A mathematical theory of communication. Bell System Technical Journal, 1948, 27(3): 379-423 |
SHI B, LIU X Z, CAO Y N, et al. Effects of salinity stress on the digestive physiology and anti-stress index of yellowtail kingfish (Seriola aureovittata). Marine Sciences, 2020, 44(6): 64-72 [史宝, 柳学周, 曹亚男, 等. 盐度胁迫对黄条 消化生理和抗应激指标的影响. 海洋科学, 2020, 44(6): 64-72] |
SLYNKO Y V, SLYNKO E E, ABLIAZOV E R, et al. Molecular- genetic identification of chameleon goby Tridentiger trigonocephalus (Gill, 1859) in the Black Sea. Ecologica Montenegrina, 2020, 32: 46-52 |
TAKASUKA A, OOZEKI Y, KUBOTA H. Multi-species regime shifts reflected in spawning temperature optima of small pelagic fish in the western North Pacific. Marine Ecology Progress Series, 2008, 360: 211-217 |
TANG Q S, SU J L, SUN S, et al. A study of marine ecosystem dynamics in the coastal ocean of China. Advances in Earth Science, 2005(12): 1288-1299 [唐启升, 苏纪兰, 孙松, 等. 中国近海生态系统动力学研究进展. 地球科学进展, 2005(12): 1288-1299] |
TENG G L, SHAN X J, JIN X S, et al. A study on the ovary-development characters and oocyte size-Distribution of Cleisthenes herzensteini in the Yellow Sea. Progress in Fishery Sciences, 2018, 39(1): 12-20 [滕广亮, 单秀娟, 金显仕, 等. 黄海高眼鲽卵巢发育特征及卵径分布. 渔业科学进展, 2018, 39(1): 12-20] |
TER BRAAK C J F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology, 1986, 67(5): 1167-1179 |
WAN R J. Morphology of early development of Sillago sihama (Forskal). Marine Fisheries Research, 1996, 17(1): 35-41 [万瑞景. 多鳞 早期发育形态. 海洋水产研究, 1996, 17(1): 35-41] |
WAN R J, JIANG Y W. Studies on the ecology of eggs and larvae of Osteichthyes in the Yellow Sea. Marine Fisheries Research, 1998, 19(1): 60-73 [万瑞景, 姜言伟. 黄海硬骨鱼类鱼卵、仔稚鱼及其生态调查研究. 海洋水产研究, 1998, 19(1): 60-73] |
WAN R J, ZHANG R Z. Fish eggs, larvae and juveniles in the offshore and adjacent waters of China. Shanghai: Shanghai Scientific and Technical Publishers, 2016 [万瑞景, 张仁斋. 中国近海及其邻近海域鱼卵与仔稚鱼. 上海: 上海科学技术出版社, 2016]
|
WAN R J, ZHAO X Y, WEI H. Spawning ecology of the anchovy Engraulis japonicus in the spawning ground of the Southern Shandong Peninsula Ⅱ. Spawning habits and embryonic developmental characteristics of the anchovy. Current Zoology, 2008, 54(6): 988-997 [万瑞景, 赵宪勇, 魏皓. 山东半岛南部产卵场鳀鱼的产卵生态Ⅱ. 鳀鱼的产卵习性和胚胎发育特性. 动物学报, 2008, 54(6): 988-997] |
WEI S. The fishing seasons and grounds of the blue spotted mackerel, Scomberomorus niphonius in the Yellwo Sea and Bohai in relation to environmental factors. Transactions of Oceanology and Limnology, 1980(2): 34-40 [韦晟. 蓝点马鲛在黄、渤海的渔场、渔期与环境的关系. 海洋湖沼通报, 1980(2): 34-40] |
WEN C N, HUANG W, CHEN K N, et al. Canonical correspondence analysisi between zooplankton community structure and environmental factors in the littoral zone of Taihu Lake. Journal of Hydroecology, 2020, 41(2): 36-44 [温超男, 黄蔚, 陈开宁, 等. 太湖滨岸带浮游动物群落结构特征与环境因子的典范对应分析. 水生态学杂志, 2020, 41(2): 36-44] |
YAN L P, ZHANG H, LI S F, et al. Progress of population identification and classification of chub mackerel (Scomber japonicus) in the East China Sea and the Yellow Sea. Marine Fisheries, 2012, 34(2): 217-221 [严利平, 张辉, 李圣法, 等. 东、黄海日本鲭种群鉴定和划分的研究进展. 海洋渔业, 2012, 34(2): 217-221] |
YU X G, DONG J, LI Y P, et al. Species composition and protection of fish eggs and larvae in the coastal waters in Liaodong Bay. Journal of Dalian Ocean University, 2018, 33(3): 370-378 [于旭光, 董婧, 李轶平, 等. 辽东湾近海鱼卵、仔稚鱼种类组成和保护分析. 大连海洋大学学报, 2018, 33(3): 370-378] |
ZHANG B. Feeding ecology of fishes in the Bohai Sea. Progress in Fishery Sciences, 2018, 39(3): 11-22 [张波. 渤海鱼类的食物关系. 渔业科学进展, 2018, 39(3): 11-22] |
ZHANG R Z, LU S F, ZHAO C Y, et al. Fish eggs and larvae in the coastal waters of China. Shanghai: Shanghai Scientific and Technical Publishers, 1985 [张仁斋, 陆穗芬, 赵 传  , 等. 中国近海鱼卵与仔鱼. 上海: 上海科学技术出版社, 1985]
, 等. 中国近海鱼卵与仔鱼. 上海: 上海科学技术出版社, 1985]
|
ZHANG X W, LIU X S. Investigation of ecology of four main commercially important marine fish species in China in the past ten years. Oceanologia et Limnologia Sinica, 1959, 2(4): 233-240 [张孝威, 刘效舜. 十年来我国四种主要海产经济鱼类生态的调查研究. 海洋与湖沼, 1959, 2(4): 233-240] |
ZHANG Y Q, QIU S Y. A preliminary study on fishery resources in Shandong coastal areas. Journal of Yantai University (Natural Science and Engineering), 2019, 32(1): 61-67 [张玉钦, 邱盛尧. 山东近海渔业资源现状初步研究. 烟台大学学报(自然科学与工程版), 2019, 32(1): 61-67] |
ZHANG Y X, BIAN X D, SHAN X J, et al. Spatio-temporal distribution of spawning grounds of chub mackerel (Scomber japonicus) and its relationship with environmental factors in the Yantai-Weihai offshore waters. Journal of Fishery Sciences of China, 2022, 29(4): 618-632 [张雨轩, 卞晓东, 单秀娟, 等. 烟威近岸海域鲐产卵场时空分布及其与环境因子的关系. 中国水产科学, 2022, 29(4): 618-632] |
ZHANG Z, XU S, CAPINHA C, et al. Using species distribution model to predict the impact of climate change on the potential distribution of Japanese whiting Sillago japonica. Ecological Indicators, 2019, 104: 333-340 |
ZHAO C Y. Marine fishery resources of China. Hangzhou: Zhejiang Science and Technology Publishing House, 1990: 106-109
|
赵 传
. 中国海洋渔业资源. 杭州: 浙江科学技术出版社, 1990: 106-109
|
ZHU S P. Investigation on Scomber japonicus fishing ground in Yantai and Weihai sea area//Expert Office of China Committee of Western Pacific Fishery Commission. Proceedings of the Second Plenary Meeting of the Western Pacific Fishery Commission. Beijing: Science Press, 1959, 1–30 [朱树屏. 烟、威海区鲐鱼渔场调查//太平洋西部渔业研究委员会中国委员会专家办公室. 太平洋西部渔业研究委员会第二次全体会议论文集. 北京: 科学出版社, 1959, 1–30]
|
ZU K W, CHENG J H, LIU Y, et al. Distribution of fishing grounds of Japanese Spanish mackerel (Scomberomorus niphonius) during spawning and overwintering period and its relation with sea surface temperature in China Sea. Transactions of Oceanology and Limnology, 2019(6): 48-57 [祖凯伟, 程家骅, 刘阳, 等. 产卵期及越冬期蓝点马鲛渔场分布变化及其与海表温度的关系. 海洋湖沼通报, 2019(6): 48-57] |